密码子是什么?关于密码子的科普介绍

密码子(codon),即信使RNA(mRNA)链翻译成何种氨基酸由三个相邻的碱基决定,亦称三联体密码。一般认为三联密码子共64个,编码20种氨基酸和1个终止密码子(stop codon)。信使RNA在细胞中能决定蛋白质分子中的氨基酸种类和排列次序。也就是说,信使RNA分子中的四种核苷酸(碱基)AUGC的序列能决定蛋白质分子中的20种氨基酸的排列顺序。转运RNA(tRNA)是密码子和氨基酸的桥梁。它一端携带氨基酸通过反密码子与密码子相结合。密码子表中一个氨基酸会对应多种密码子。这种现象称为密码子的简并性,目的是为了提高翻译的容错率。绝大多数起始密码子为AUG,部分原核生物以GUG/UUG为起始密码子。终止密码子为UAA 、UGA和UAG。密码子的发现是上个世纪分子生物学最重要的发现之一。它的发现历经数理推断和实验验证的阶段。
密码子定义
科学家把信使RNA链上决定一个氨基酸的相邻的三个碱基叫做一个“密码子”,亦称三联体密码。
构成RNA的碱基有四种,分别为AUGC。从理论上分析碱基的组合有4的3次方=64种,64种碱基的组合即64种密码子。仔细分析20种氨基酸的密码子表,就可以发现,同一种氨基酸可以由几个不同的密码子来决定,起始密码子为AUG(甲硫氨酸) , 另外还有UAA(赭石)、UAG(琥珀)、UGA(蛋白石)三个密码子不能决定任何氨基酸,是蛋白质合成的终止密码子。AUG在原核生物中翻译成甲酰甲硫氨酸,在真核生物中翻译成甲硫氨酸。
遗传信息、密码子、反密码子的区别与联系:
遗传信息是指DNA分子中基因上的脱氧核苷(碱基)排列顺序,密码子是指信使RNA上决定一个氨基酸的三个相邻碱基的排列顺序,反密码子(anticodon)是指转运RNA上的一端的三个碱基排列顺序。其联系是:DNA(基因)的遗传信息通过转录传递到信使RNA上,转运RNA一端携带氨基酸,另一端反密码子与信使RNA上的密码子(碱基)配对。
密码子起源
除了少数的不同之外,地球上已知生物的遗传密码均非常接近;因此根据演化论,遗传密码应在生命历史中很早期就出现。现有的证据表明遗传密码的设定并非是随机的结果,对此有以下的可能解释:
一项研究显示,一些氨基酸与它们相对应的密码子有选择性的化学结合力,这显示复杂的蛋白质制造过程可能并非一早存在,最初的蛋白质可能是直接在核酸上形成。 原始的遗传密码可能比今天简单得多,随着生命演化制造出新的氨基酸再被利用而令遗传密码变得复杂。虽然不少证据证明这观点,但详细的演化过程仍在探索之中,。 经过自然选择,现时的遗传密码减低了突变造成的不良影响。一般分子生物学教材中介绍的密码子进化学说为Dayhoff假说。Dayhoff及其同事认为密码子的进化路线为GNC→GNY→RNY→RNN→NNN。其中Y代表C或U两种嘧啶,R代表G或A两种嘌呤。在GNC和GNY阶段,第二位碱基决定氨基酸种类;RNY阶段前两位碱基决定氨基酸种类;RNN和NNN阶段三位碱基全部参与氨基酸种类决定。
密码子特点
①. 遗传密码子是三联体密码:一个密码子由信使核糖核酸(mRNA)上相邻的三个碱基组成。
② 密码子具有通用性:不同的生物密码子基本相同,即共用一套密码子。
③ 遗传密码子无间隔:两个密码子间没有间隔,密码子与密码子之间没有任何不编码的核苷酸,读码必须按照一定的读码框架,从正确的起点开始,一个不漏地一直读到终止信号。
④ 遗传密码子不重叠:在多核苷酸链上任何两个相邻的密码子不共用任何核苷酸。
⑤ 密码子具有简并性:除了甲硫氨酸和色氨酸外,每一个氨基酸都至少有两个密码子。这样可以在一定程度内,使氨基酸序列不会因为某一个碱基被意外替换而导致氨基酸错误。
⑥ 密码子阅读与翻译具有一定的方向性:从5'端到3'端。
⑦有起始密码子和终止密码子:起始密码子有两种,一种是甲硫氨酸(AUG),一种是缬氨酸(GUG),而终止密码子(有3个,分别是UAA、UAG、UGA)没有相应的转运核糖核酸(tRNA)存在,只供释放因子识别来实现翻译的终止。
在信使RNA中,碱基代码A代表腺嘌呤,G代表鸟嘌呤,C代表胞嘧啶,U代表尿嘧啶(注意:RNA与DNA不同,RNA没有胸腺嘧啶T,取而代之的是尿嘧啶U,按照碱基互补配对原则,U与A形成配对)。
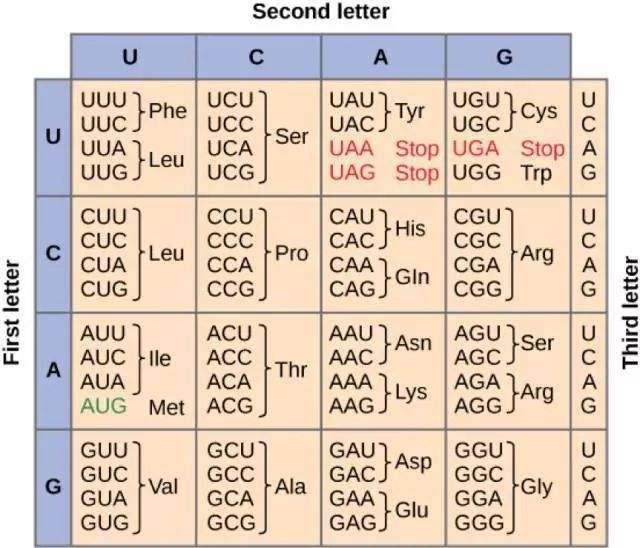
密码子表
第一位碱基A
AUU (Ile/I)异亮氨酸 AUC (Ile/I)异亮氨酸 AUA (Ile/I)异亮氨酸 AUG (Met/M)甲硫氨酸(起始)
ACU (Thr/T)苏氨酸 ACC (Thr/T)苏氨酸 ACA (Thr/T)苏氨酸 ACG (Thr/T)苏氨酸
AAU (Asn/N)天冬酰胺 AAC (Asn/N)天冬酰胺 AAA (Lys/K)赖氨酸 AAG (Lys/K)赖氨酸
AGU (Ser/S)丝氨酸 AGC (Ser/S)丝氨酸 AGA (Arg/R)精氨酸 AGG (Arg/R)精氨酸
第 一 位 碱 基 U
UUU (Phe/F)苯丙氨酸 UUC (Phe/F)苯丙氨酸 UUA (Leu/L)亮氨酸 UUG (Leu/L)亮氨酸
UCU (Ser/S)丝氨酸 UCC (Ser/S)丝氨酸 UCA (Ser/S)丝氨酸 UCG (Ser/S)丝氨酸
UAU (Tyr/Y)酪氨酸 UAC (Tyr/Y)酪氨酸 UAA (终止) UAG (终止)
UGU (Cys/C)半胱氨酸 UGC (Cys/C)半胱氨酸 UGA (终止) UGG (Trp/W)色氨酸
第 一 位 碱 基 C
CUU (Leu/L)亮氨酸 CUC (Leu/L)亮氨酸 CUA (Leu/L)亮氨酸 CUG (Leu/L)亮氨酸
CCU (Pro/P)脯氨酸 CCC (Pro/P)脯氨酸 CCA (Pro/P)脯氨酸 CCG (Pro/P)脯氨酸
CAU (His/H)组氨酸 CAC (His/H)组氨酸 CAA (Gln/Q)谷氨酰胺 CAG (Gln/Q)谷氨酰胺
CGU (Arg/R)精氨酸 CGC (Arg/R)精氨酸 CGA (Arg/R)精氨酸 CGG (Arg/R)精氨酸
第 一 位 碱 基 G
GUU (Val/V)缬氨酸 GUC (Val/V)缬氨酸 GUA (Val/V)缬氨酸 GCU (Ala/A)丙氨酸
GCC (Ala/A)丙氨酸 GAU (Asp/D)天冬氨酸 GAC (Asp/D)天冬氨酸 GAA (Glu/E)谷氨酸
GGU (Gly/G)甘氨酸 GGC (Gly/G)甘氨酸 GCA (Ala/ A)丙氨酸 GCG (Ala/A)丙氨酸
GUG (Val/V)缬氨酸 GAG (Glu/E)谷氨酸 GGG (Gly/G)甘氨酸 GGA (Gly/G)甘氨酸
研究历程
基因密码的破译是六十年代分子生物学最辉煌的成就。先后经历了五十年代的数学推理阶段和1961-1965年的实验研究阶段。1954年,物理学家George Gamov根据在DNA中存在四种氨基酸,在蛋白质中存在二十种氨基酸的对应关系,做出如下数学推理:如果每一个核苷酸为一个氨基酸编码,只能决定四种氨基酸;如果每二个核苷酸为一个氨基酸编码,可决定16种氨基酸(42=16)。上述二种情况编码的氨基酸数小于20种氨基酸,显然是不可能的。那么如果三个核苷酸为一个氨基酸编码的,可编64种氨基酸(43=64);若四个核苷酸编码一个氨基酸,可编码256种氨基酸(44=256),以此类推。Gamov认为只有4^3=64这种关系是理想的,因为在有四种核苷酸条件下,64是能满足于20种氨基酸编码的最小数。而44=256以上。虽能保证20种氨基酸编码,但不符合生物体在亿万年进化过程中形成的和遵循的经济原则,因此认为四个以上核苷酸决定一个氨基酸也是不可能的。1961年,Brenner和Grick根据DNA链与蛋白质链的共线性(colinearity),首先肯定了三个核苷酸的推理。随后的实验研究证明上述假想是正确的。
遗传密码的破译实在体外蛋白质合成体系建立和核酸人工合成的技术基础上成长起来的。值得一提的是在破译密码的竞赛中,美国的尼伦伯格博士走在前面。主要的方法有两种:
-
以均聚物和随机共聚物和特定序列的共聚物为模板指导多肽的合成。在体外无细胞蛋白质合成体系中加入人工合成的polyU 开创了破译遗传密码的先河。自1961年发现mRNA后,许多实验室开始在无细胞蛋白质合成系统中加入mRNA,去研究蛋白质生物合成过程,并表明加入mRNA能刺激无细胞系统中蛋白质合成。1961年,美国NIH的Nirenberg和Mathaei,设想:即然mRNA有刺激无细胞系统中的蛋白质合成作用,加入人工合成的多聚核苷酸亦将会有这种促进作用。按此设想,他们合成了polyU作为模板,以观察无细胞系统中蛋白质合成速率。因为在反应体系中加入高Mg2+浓度,可有利于IF(起始因子)的作用和fMet-tRNANet的形成,从而保证肽链合成的起始不需mRNA的适当信号。当把翻译产物分离、纯化和做序列分析后,结果出乎意料,合成的肽链中的氨基酸残基全部是苯丙氨酸,即polyPhe。于是第一次确认了UUU是Phe的密码子。这样,就在一个偶然的机会开创了破译密码的工作。随后,他们又以polyA和polyC为模板,证明了分别可指导合成polyLys和polyPro,即确定了AAA是Lys的密码子,CCC是pro的密码子。
-
混合共聚物(mixed copolymers)实验对密码子中碱基组成的测定。1963年,Speyer和Ochoa等发展了用两个碱基的共聚物破译密码的方法。例如,以A和C原料,合成polyAC。polyAC含有8种不同的密码子:CCC、CCA、CAA、AAA、AAC、ACC、ACA和CAC。各种密码子占的比例随着A和C的不同而不同,例如当A和C的比例等于5:1时,AAA:AAC的比例=5× 5× 5:5× 5× 1=125:25。依次类推。实验显示AC共聚物作模板翻译出的肽链由六种氨基酸组成,它们是Asp,His,Thr,Pro,和Lys,其中Pro和Lys的密码子早先已证明分别是CCC和AAA。根据共聚物成份不同的比例和翻译产物中氨基酸比例亦不同的关系,Speyer等确定了Asp、Glu和Thr的密码子含2AlC;His的密码子含1A2C;Thr的密码子也可以含1A2C;Pro为3C或1A2C;Lys为3A。但上述方法不能确定A和C的排列方式,而只能显示密码子中碱基组成及组成比例。例如,Asp,Glu和Thr的2A1C可能有三种排列方式,即AAC、ACA、CAA。此外,通过反复改变共聚物成份比例的方法亦十分麻烦和费时。
-
用重复共聚物(repeating copolymers)破译密码。几乎在上述Nirenberg和Leder工作的同时,Nishimura,Jones,和Khorana等人应用有机化学和酶学技术,制备了已知的核苷酸重复序列。蛋白质在核糖体上的合成可以在这些有规律的共聚物的任一点开始,并把特异的氨基酸参入肽链。例如,重复序列CUCUCUCUCU......是多肽Leu-Ser-Leu-Ser......或者是多肽Ser-Leu-Ser......的信使分子.使用共聚物构成三核苷酸为单位的重复顺序,如(AAG)n,它可合成三种类型的多肽:polyLys、polyArg和polyGlu,即AAG是Lys的密码子,AGA是Arg的密码子,GAA是Glu的密码子。又如(AUC)n序列是polyIle、polySer和polyHis的模板。
-
2、aa-tRNA与确定的三核苷酸序列结合。正当Speyer等人按上述方法奋力时,Nirenberg和Leder于1964年建立了破译密码的新方法,即tRNA与确定密码子结合实验。该方法利用了如下事实:即是在缺乏蛋白质合成所需的因子的条件下,特异氨基酸-tRNA(aa-tRNA)也能与核糖体-mRNA复合物结合。最重要的是这种结合并不一定需要长的mRNA分子,而三核苷酸实际上就可以与核糖体结合。例如,当polyU与核糖体混合时,仅有Phe-tRNA(苯丙氨酰-tRNA)与之结合;相应地Pro-tRNA(脯氨酰-tRNA)特异地与polyC结合。还有GUU可促进Val-tRNA(缬氨酰-tRNA)结合,UUG促进Leu-tRNA(亮氨酰-tRNA)结合等。虽然所有64个三核苷酸(密码子)都可按设想的序列合成,但并不是全部密码子均能以这种方法决定因为有一些三核苷酸序列与核糖体结合并不象UUU或GUU等那样有效,以致不能确定它们是否能为特异的氨基酸编码。
如此至1965年破译了所有氨基酸的密码子。
伴随着实验技术和生物理论的发展,三联体密码并非一成不变的。例如,2014年Scripps研究所的Floyd Romesberg在AUGC外又引入两个人工碱基X和Y,并设计了专门的tRNA。最终大肠杆菌成功表达了引入的人工碱基。某种意义上密码子被扩展到六碱基水平。
2019年5月16日,顶尖学术期刊《Nature》上线的一篇论文载:英国剑桥MRC分子生物学实验室的Jason Chin教授与其同事就在大肠杆菌中实现了目标。他们重新编码了一个大肠杆菌菌株的全部基因组,只用59个密码子就合成出所有的必需氨基酸,代表终止信号的密码子也从3个压缩为2个。而“节省”下来的密码子,可以为将来在活细胞内生成非天然的“定制蛋白质”提供合成空间。
本文转载于:搜狗科学,baike.sogou.com/kexue/d19764594964702473.htm,本内容使用CC-BY-SA 3.0授权,用户转载请注明出处





















