生态位是什么?关于生态位的科普介绍

在生态学中,生态位是一个物种与特定环境条件的匹配。它描述了一个生物体或群体如何对资源和竞争对手的分布做出响应(例如,当资源丰富时,当捕食者、寄生虫和病原体稀缺时,通过生长),以及它如何反过来改变这些相同的因素(例如,限制其他生物体对资源的获取,充当捕食者和猎物消费者的食物来源)。“构成环境生态位维度的变量的类型和数量因物种而异,特定环境变量对一个物种的相对重要性可能因地理和生物环境而异”。
格林尼利亚生态位由物种生活的栖息地及其伴随的行为适应决定。埃尔顿生态位强调,一个物种不仅生长在环境中并对环境做出反应,它还可能随着生长而改变环境及其行为。哈钦森生态位使用数学和统计来试图解释物种如何在给定的社区中共存。
生态位的概念是生态生物地理学的核心,它关注生态群落的空间格局。“物种分布及其随时间的动态是由物种特性、环境变化...、以及两者之间的相互作用——特别是一些物种,尤其是我们自己的物种,改变它们的环境和改变许多其他物种的范围动态的能力。”居民改变生态位的主题是生态位建设。
大多数物种存在于标准生态位中,与同一大类分类学中的其他密切相关的物种相似,具有共同的行为、适应性和功能特征,但也有例外。非标准生态位填充物种的一个首要例子是新西兰的不会飞的、居住在地面的猕猴桃鸟,它以蠕虫和其他地面生物为食,生活在类似哺乳动物的生态位中。岛屿生物地理学可以帮助解释岛屿物种和相关的未填充生态位。

格林内利生态位

生态位的生态意义来自于生态位作为雕像墙中凹处的意义,它本身可能来源于中古法语这个词nicher,意思是筑巢。这个术语是自然学家罗斯韦尔·希尔·约翰逊创造的但是约瑟夫·格林内尔在1917年的论文《加州付令超的生态位关系》中可能是第一个在研究项目中使用它的人。
格林尼利亚生态位概念体现了这样一种观点,即物种的生态位由它生活的栖息地及其伴随的行为适应决定。换句话说,生态位是允许物种持续存在并产生后代的栖息地要求和行为的总和。例如,加州打谷机的行为与其生活的夏帕拉尔栖息地一致——它在灌木丛中繁殖和进食,并通过从灌木丛拖到灌木丛中逃离捕食者。它的“生态位”是由这个栖息地恰当地补充了打鱼的行为和身体特征(伪装的颜色、短翅膀、强壮的腿)来定义的。
生态位的这种观点允许生态等同物和空生态位的存在。生态等值种生物是来自不同分类组的生物,在相似的生境中表现出相似的适应性,例如在美国和非洲沙漠中发现的不同的肉质植物,仙人掌和大戟。作为另一个例子,大安的列斯群岛的变色龙蜥蜴是趋同进化、适应性辐射和生态等价物存在的罕见例子:变色龙在彼此独立的相似的微生境中进化,并在所有四个岛屿上产生相同的生态变体。
埃尔顿生态位
1927年,查尔斯·萨瑟兰·埃尔顿,一个英国人 生态学家定义生态位如下:“动物的‘生态位’意味着它在生物环境的位置,它与食物和敌人的关系。"
埃尔顿根据觅食活动(“食物习惯”)对生态位进行了分类:
For instance there is the niche that is filled by birds of prey which eat small animals such as shrews and mice. In an oak wood this niche is filled by tawny owls, while in the open grassland it is occupied by kestrels. The existence of this carnivore niche is dependent on the further fact that mice form a definite herbivore niche in many different associations, although the actual species of mice may be quite different.
从概念上来说,埃尔顿生态位引入了物种的概念反应 到和对...的影响环境。与其他生态位概念不同,它强调物种不仅生长在基于可用资源、捕食者和气候条件的环境中并对其做出响应,还随着生长改变这些因素的可用性和行为。在一个极端的例子中,海狸需要某些资源来生存和繁殖,但也需要建造大坝来改变海狸生活的河流中的水流。因此,海狸影响生活在流域内和附近的其他物种的生物和非生物条件。在一个更微妙的情况下,以不同速度消耗资源的竞争对手可能导致不同物种之间资源密度的循环。物种不仅在资源密度方面增长不同,而且它们自己的种群增长也会随着时间的推移影响资源密度。
哈钦森生态位

哈钦森生态位是一个“ n维超体积”,其中维是环境条件和资源,定义了个人或物种实践“其”生活方式的要求,更具体地说,是对其种群持续存在的要求。“超体积”定义了资源的多维空间(例如光、营养物、结构等)。)对于生物体是可用的(并且被生物体特别使用),并且“除了那些被考虑的物种之外的所有物种都被认为是坐标系的一部分”。
生态位概念于1957年由动物学家乔治·伊夫林·哈钦森推广。哈钦森探究了为什么在任何一个栖息地都有这么多种生物的问题。他的工作启发了许多其他人开发模型来解释给定群落中共存物种的数量和相似程度,并导致了“生态位宽度”(给定物种使用的资源或生境的多样性)、“生态位划分”(共存物种的资源分化)和“生态位重叠”(不同物种使用资源的重叠)的概念。
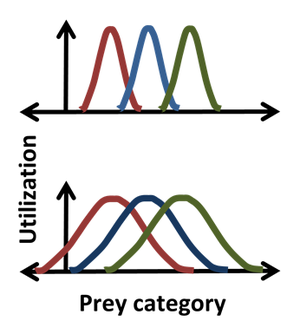
罗伯特·麦克阿瑟和理查德·莱文斯利用采用直方图描述作为哈钦森坐标函数的“发生频率”的“资源利用”生态位,将统计学引入哈钦森生态位。因此,例如,高斯可以描述一个物种吃特定大小猎物的频率,给出比简单指定一些中值或平均猎物大小更详细的生态位描述。对于这样的钟形分布位置,宽度和形式生态位对应于中间,标准偏差和实际分布本身。图中显示了使用统计数据的一个优势,其中很明显,对于较窄的分布(顶部),极左和极右物种之间不存在对猎物的竞争,而对于较宽的分布(底部),生态位重叠表明所有物种之间都可能发生竞争。资源利用方法在于假设不仅竞争能发生,而且它做且资源利用中重叠直接使得能够估计竞争系数。然而,这一假设可能会被误导,因为它忽略了每个类别的资源对生物体的影响以及生物体对每个类别的资源的影响。例如,重叠区域中的资源可以是非限制性的,在这种情况下,尽管生态位重叠,也不存在对该资源的竞争。
一个不受其他物种干扰的生物体可以利用它能够生存和繁殖的所有条件(生物和非生物)和资源,这被称为它的基础生态位。然而,由于来自其他生物的压力和与其他生物的相互作用(即相互特异性竞争),物种通常被迫占据比这更窄的生态位,并且它们大多高度依赖于这一生态位适合的;这被称为实际生态位。哈钦森使用资源竞争的概念作为驱动生态的主要机制,但过分强调这一重点已被证明是生态位概念的障碍。特别是,过分强调一个物种对资源的依赖,导致对生物体对其环境的影响,例如定居和入侵的重视太少。
术语“适应区”是古生物学家乔治·盖洛德·辛普森创造的,用来解释一个种群如何从一个生态位跳到另一个适合它的生态位,跳到一个“适应区”,这个适应区是通过一些修改,或者可能是食物链的变化实现的,这使得适应区对它来说是可用的,而它的生活方式没有中断,因为这个群体“预先适应”了新的生态机遇。

哈钦森的“生态位”(对一个物种占据的生态空间的描述)与格林内尔定义的“生态位”略有不同(一个生态角色,实际上可能由一个物种占据,也可能不由一个物种占据——见空白壁龛)中。
生态位是由单一物种占据的生态空间中非常特定的一部分。假设没有两个物种在所有方面是相同的(称为哈定的“不平等公理”))和竞争排斥原理,一些资源或适应性维度将提供每个物种特有的生态位。然而,物种可以共享一种“生活方式”或“生态策略”,这是生态空间的更广泛定义。例如,澳大利亚草原物种,虽然不同于大平原草原,但表现出相似的生活方式。
一旦生态位被空置,其他生物可以填补这个位置。例如,因塔盘灭绝而留下的生态位被其他动物(特别是小马品种科尼克)填补了。此外,当植物和动物被引入新的环境时,它们有可能占据或侵入土著生物的生态位,通常超过土著物种。人类将非本土物种引入非本土栖息地通常会导致外来物种或入侵物种的生物污染。
物种在生态空间中基本生态位的数学表示,及其随后向地理空间的投影,是生态位建模的领域。
因素
不同的维度,或者绘图轴生态位代表不同的生物和非生物变量。这些因素可能包括生物体的生活史、栖息地、营养位置(位于食物链中)和地理范围的描述。根据竞争排斥原理,没有两个物种可以在同一环境中长时间占据同一生态位。实际生态位的参数由该物种的实际生态位宽度来描述。一些被称为专家的植物和动物需要特定的栖息地和环境才能生存,例如专门生活在古老森林中的斑点猫头鹰。其他被称为多面手的植物和动物并不特别,它们可以在一系列条件下生存,例如蒲公英。
本文转载于:搜狗科学,baike.sogou.com/kexue/d11072.htm,本内容使用CC-BY-SA 3.0授权,用户转载请注明出处
下一篇:杂交是什么?关于杂交的科普介绍