恐龙类是什么?关于恐龙类的科普介绍

恐龙是恐龙总目(学名:Dinosauria)的通称,是生活在大约2亿3500万年至6500万年前、能以后肢支撑身体直立行走的多样化优势陆栖脊椎动物,支配全球陆地生态系统超过1亿6千万年之久。所有非鸟恐龙以及大部分鸟形恐龙所在白垩纪末(约7000万-6800万年前)灭绝事件中绝灭(即俗称的恐龙大灭绝)大部分恐龙已经灭绝,仅有鸟形恐龙中的鸟类存活下来并且繁荣至今。
恐龙在6500万年前白垩纪结束的时候突然全部消失,成为地球生物进化史上的一个谜,这个谜仍无人能解。地球过去的生物,均被记录在化石之中。中生代的地层中,即曾发现许多恐龙的化石。其中可以见到大量或呈现各式各样形状的骨骼。但是,在紧接着的新生代地层中,却完全看不到非鸟恐龙的化石,由此推知非鸟恐龙在中生代时一起灭绝了,如今仅存鸟类,大多数科学家都认可“鸟类属于恐龙”的说法。
恐龙种类很多,体形和习性相差也大,其中最大的易碎双腔龙可能超过50米,而最小的蜂鸟可能还不到10厘米。就食性来说,有温顺的草食者和凶暴的肉食者,还有荤素都吃的杂食性恐龙。统治了地球大约8000万年(1.44亿年前---6500万年前)
介绍
现可追溯年代最久远的恐龙化石是在1677年,由当时英国牛津大学博物学家Robert Plot (罗伯特‧普劳特)发现的恐龙化石,这批化石后于1824年由英国地质学家 威廉‧布克兰命名为“斑龙”。
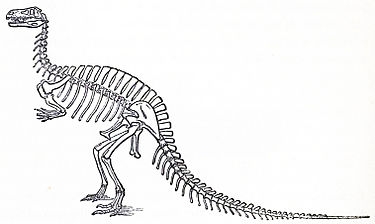
恐龙(Dinosauria)是在1842年由英国古生物学家理查·欧文正式提出,欧文把恐龙总目定义为蜥形纲的一个独特种类或亚目,该目包含当时在英国及世界各地发现的恐龙。
根据系统发生学的定义,恐龙包含了腕龙、三角龙与现代鸟类最近的共同祖先,以及它们最近共同祖先的所有后代。根据理查·欧文的定义,恐龙被定义为斑龙、林龙与禽龙这三个属的最近共同祖先。这两个定义的范围相近,都包含兽脚亚目、蜥脚形亚目、剑龙亚目、甲龙亚目、角龙亚目、厚头龙亚目、以及鸟脚亚目等演化支。早在近年种系发生学研究盛行前,上述分类单元皆已在科学文献中出现、使用,并持续到今日。
现今古生物学界普遍同意鸟类是恐龙的直系后代。根据严格的亲缘分支分类法定义,一个天然的演化支应包含单一共同祖先的所有后代。若以此方法来看,所有鸟类都为恐龙,而恐龙并没有灭亡。大部分古生物学家将鸟类归类于恐龙-蜥臀目-兽脚亚目-虚骨龙类-手盗龙类。
恐龙开始时被列为爬行类的一目,但现是独立的一纲。根据其骨盘构造的不同,可区分为蜥臀目(saurischia)和鸟臀目(Ornithischia)两目。过去所说的禽龙是根据R·欧文(R.Owen 1912年)命名的。在中生代从三叠纪到白垩纪非常繁盛,中生代末新生代初绝灭。曾分布于6个大陆,适应各种环境,有肉食性的和草食性的。前肢比后肢短,多数只用后肢步行,但其中也有四肢步行的。侏罗纪以后出现了颈尾均长的巨大的类型(载域龙,Atloantosaurus 体长30米以上,体重约50吨),牙齿深埋于齿槽内,并局限在颚骨的周缘。脊椎骨有双凹型、平凹型、后四型的区别,但其后代几乎都为后凹型。有的种类与鸟相近,骨中空,如霸王龙(Tyrranosaurus),雷龙(Brontosaurus),禽龙(Lguanodon),剑龙(Stegosaurus),钩龙(Ankylosaurus),三角龙(Triceratops)等都是。在南库页岛产的日本龙(Nipponosaurus)是属于草食龙,是鸭嘴龙类之一种。日本长畸县高岛煤矿内有类似的小残片发现。最近的化石研究发现了一种新型驰龙(Dineobellator notohesperus),驰龙是生活在白垩纪时期的一种身覆羽毛的中小型肉食性恐龙,此次的化石研究发现增进了我们对白垩纪时期的恐龙演化的理解。
鉴定特征
近年来的新发现化石,使得科学家们越来越难列出一致性的恐龙鉴定特征。恐龙的骨骼具有一些演化适应,以此将它们与原始的主龙类祖先辨别出来。在后期的恐龙中,这些特征经过更多的演化改变。
恐龙的共同特征:
肠骨后部有个突出区块。
胫骨末端边缘宽广,有个往后的凸缘。
肱骨有低矮的三角嵴(Deltopectoral crest,让胸锁三角肌肉附着的部分),长度约是肱骨的1/3到1/2。
距骨有个明显上突,与胫骨契合。
除上述几个特征以外,大部分恐龙还有一些共同特征,但是因为出现在其他主龙类,或不存在于早期的恐龙身上,因此不列为恐龙的共有衍征。举例而言,恐龙具有两对颞颥([niè rú])孔,这是双孔动物的共同特征,恐龙还具有眶前孔与下颌孔,这些则是主龙类的共同特征。另外几个特征,早在恐龙之前已经出现,消失于最早期的恐龙,而在不同的恐龙生物群独自演化出现,包含:延长的肩胛骨、至少三个愈合的荐椎(其他主龙类也有三个愈合荐椎,艾雷拉龙只有两个愈合荐椎)、开放式有孔的髋([kuān])臼(农神龙的髋臼闭合)。恐龙的鉴定特征难以确认,部分原因则是三叠纪晚期的恐龙与同时期其他主龙类的化石不多,两者却有许多相似处,使得某些主龙类曾被误认为是恐龙。
恐龙具有直立的步态,类似大部分的现代哺乳类,而大部分其他爬行动物则是四肢往两侧延展的步态。恐龙的臀窝朝向两侧,股骨的第四粗隆部往内侧,两者契合,产生直立的步态。直立的步态可让恐龙快速运动时,可以同时呼吸,使它们成为活跃的动物,较采取两侧延展步态的动物更具优势。直立步态可以降低四肢弯曲时所承受的压力,有助于恐龙发展出巨大的体型。
起源
恐龙在过去被认为是复系群,包含数个毫不相干的演化支,并非来自于共同的最近祖先,因为身处类似的生态环境中,而个别演化出类似的形态与特征。自从70年代的恐龙文艺复兴以来,恐龙已被认为是单系群,具有最近的共同祖先,是个天然演化支。
恐龙与它们的主龙类祖先在三叠纪中到晚期分开演化,接近2亿3000万年前,大约是二叠纪-三叠纪灭绝事件后约2000万年,该次灭绝事件消灭了接近95%的地球生物物种。对于早期恐龙始盗龙发现地层的放射线定年法检验,证实始盗龙生存于三叠纪晚期,古生物学家认为始盗龙的外表类似所有恐龙的共同祖先;如果属实,则显示最早的恐龙应该是种小型、二足掠食动物。在阿根廷的三叠纪中期地层所发现的原始、类似恐龙的鸟颈类主龙,例如:马拉鳄龙与兔蜥,也支持这个说法;已出土的化石证据显示这群动物都为小型、二足掠食动物。
恐龙刚出现时,陆地上有多种不同的动物,例如坚蜥目、劳氏鳄类、鸟鳄科、喙头龙目等原始主龙形下纲,以及二齿兽下目与犬齿兽亚目等兽孔目动物。这些动物大部分在三叠纪后期的两次灭绝事件中灭亡。第一次灭绝事件发生于卡尼阶与诺利阶交接处,许多原始主龙形下纲动物在此事件中灭亡,例如原龙目与喙头龙目。第二次灭绝事件是三叠纪-侏罗纪灭绝事件,发生于约2亿年前,二齿兽类与大部分的早期主龙类灭亡,包含:坚蜥类、鸟鳄科、植龙目、与劳氏鳄类。这群动物灭亡后,鳄形超目、恐龙、哺乳类、翼龙目、有鳞目与乌龟开始占据陆地的生态系统。
原始恐龙的第一次适应演化发生于三叠纪的卡尼阶与诺利阶之间,极可能因为当时有许多优势动物灭亡。在传统的观念里,在三叠纪中期,恐龙与当时其他的陆地动物展开长时间的竞争,并取得优势,但这是不可能的。根据化石纪录,恐龙没那么早出现,且早期恐龙并非稳定的增加、多样化,而是后来才急骤的大量演化;如果它们与其他动物有过长时间竞争,应该会在化石纪录中反映出来。恐龙在卡尼阶时的数量非常稀少,仅占陆地动物群的1~2%。在诺利阶后,数个动物群灭亡后,恐龙则占了陆地动物群的50~90%。恐龙的直立步态过去被视为成功演化的关键,但现在已知有数个同时代的动物群也有直立步态,例如:坚蜥目、鸟鳄科、劳氏鳄类、以及数种鳄形超目动物。三叠纪后期是个生物剧变的时代,有许多植物、海生动物兴起衰落,三叠纪后期的气候也发生巨大变化。
种类概要

许多的古生物学家相信恐龙形成单一的动物族群,即恐龙类(Dinosauria)。它们与爬行动物最大区别在於它们的站立姿态与行进方式。恐龙具有全然直立的姿势,其四肢构建在其体躯的正下方位置。这样的架构要比那些其它种类的爬行动物,像是鳄类(其四肢向外侧伸展方式),在走路与奔跑上更有效益,更为方便。恐龙和其它灭绝的爬行动物能够明显的区分,主要是根据其腰带(骨盘构造)与四肢的骨骼学特征,这些在演化上的发展,能够让恐龙得以站立奔走。

恐龙指的是特定陆地爬行动物,能采取直立步态,在经典分类中包括蜥臀目(像蜥蜴似的髋骨)与鸟臀目(像鸟类的髋骨),并不包括翼龙目、鱼龙目、蛇颈龙目、沧龙科。恐龙常被认为是总目,或是未定位的演化支。蜥臀目和鸟臀目以其骨盆(髋骨)结构来区分。
在蜥臀类中(学名:Saurischia),其骨盆非常类似于现代的爬行动物,其髋骨系统中,耻骨伸前前方,而坐骨则伸向体躯的後方。蜥臀目主要包括两个亚目:二足、大
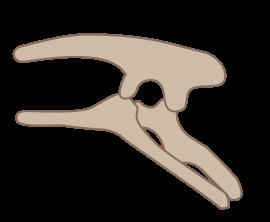
在鸟臀类中(学名:Ornithischia),则具有较为复杂的腰带构造。其中髋骨系统中,耻骨已经转向体躯的后方,而与坐骨呈平行排列,同时鸟臀类在下颌骨也发展出了前齿骨,这是趋同演化的结果。与鸟类不同的是,鸟臀目的耻骨有个朝前的突出部分。鸟臀目包含多样化的植食性动物。
根据传统定义,蜥臀目的范围是:与鸟类具有最近共同祖先,而与鸟臀目具有较远共同祖先的所有恐龙;鸟臀目的范围则是:与三角龙具有最近共同祖先,而与蜥臀目具有较远共同祖先的所有恐龙。
种属名录恐龙分类表
| 蜥臀目 | 兽脚亚目 | 坚尾龙类 | 斑龙超科:皮亚尼兹基龙科、斑龙科、棘龙科 肉食龙类:顶棘龙属、异特龙超科 虚骨龙类:美颌龙科、暴龙超科、手盗龙形类 | |
| 角鼻龙类 | 新角鼻龙类:角鼻龙科、阿贝力龙超科 | |||
| 蜥脚形亚目 | 早期蜥脚类 | 瓜巴龙科、板龙科、里奥哈龙科、大椎龙科、近蜥龙类 | ||
| 蜥脚下目 | 火山齿龙科、鲸龙科、马门溪龙科、图里亚龙类 | |||
| 新蜥脚类 | 梁龙超科:雷巴齐斯龙科、叉龙科、梁龙科 | |||
| 大鼻龙类:圆顶龙科、腕龙科、盘足龙科、泰坦巨龙类 | ||||
| 鸟臀目 | 装甲类 | 腿龙科、剑龙亚目、甲龙亚目 | ||
| 角足龙类 | 鸟脚亚目 | 薄板类、腱龙科、凹齿龙科、橡树龙科 鸭嘴龙形类:禽龙科、鸭嘴龙超科 | ||
| 头饰龙类 | 厚头龙亚目、角龙亚目 | |||
生活习性
对恐龙的研究来自于多样化的非化石与化石纪录,例如:粪化石、化石、足迹化石、羽毛、胃石、皮肤痕迹、内脏、与软组织。物理学(尤其是生物力学)、生物学、化学、地球科学等相关学科的交叉应用研究恐龙相关知识。恐龙的体型与行为是其中最重要的研究方向。
行为
对恐龙行为的假设,多是根据化石的姿态、栖息地、电脑模拟的生物力学、以及与类似生态位的现代动物比较而来。有些行为假设存在争议,目前科学家们普遍同意鳄鱼与鸟类所共同拥有的行为,有可能也存在于它们的近亲非鸟恐龙身上。
在1878年,比利时的贝尼沙特发现了31个禽龙化石,可能是洪水所造成的集体死亡,这是恐龙群居行为的第一个证据,之后在其他的地层也发现集体死亡的现象,再加上群集的足迹化石,显示许多恐龙物种具有群居的行为。目前已发现由数百个、甚至数千个个体所留下的鸭嘴龙科足迹化石,显示鸭嘴龙科可能会大量群体性移动,如同今日的美洲野牛与非洲跳羚。在英格兰牛津发现的蜥脚类恐龙的足迹化石,显示不同的蜥脚类恐龙会集体性移动。恐龙群体行动可能有许多原因,例如防御掠食动物、集体迁徙,或者保护幼年个体。举例而言,中国内蒙古曾发现20多个群聚的中国似鸟龙化石,年龄为1到7岁,科学家认为这些中国似鸟龙是在集体行动时,陷在泥泞中死亡。
在1978年,古生物学家杰克·霍纳(Jack Horner)于蒙大拿州发现慈母龙的化石与蛋巢遗迹,显示鸟脚亚目具有长时间的亲代养育行为。
在1997年,在巴塔哥尼亚白垩纪地层发现的泰坦巨龙类化石,显示这些大型恐龙也具有筑巢的行为。
某些恐龙具有头冠或是头盾,例如头饰龙类、兽脚亚目、以及赖氏龙亚科,它们的头冠、头盾过于脆弱,无法用来自我防卫,而可能具有性展示功能,或是使用在物种内打斗行为,但是目前对于恐龙的求偶与领域性行为所知有限,故尚不能确定。一些兽脚亚目化石的头部具有伤口,可能是同类互相攻击的后果。恐龙的沟通方式仍充满谜题,是个正在研究中的领域。近年的研究显示,赖氏龙亚科的中空头冠可能具有扬声器作用,可用来与同类之间做声音的沟通、辨认。
根据目前的化石证据,某些植食性恐龙可能是穴居动物,例如棱齿龙类的掘奔龙;而某些兽脚类恐龙则是树栖动物,例如原始的驰龙科小盗龙。但大部分恐龙则是依靠陆地为生存。生物力学可以用来研究恐龙的移动方式,进而研究它们的相关行为模式。借由计算恐龙的肌肉力量传递模式,以及重心位置,可推算出恐龙的移动速度。
体型
尽管目前证据不全,但是恐龙的体型的上限很大,当然这是因为当时地球环境保持长时间的稳定与相对温暖的缘故,而非现在的人类所处的冰河期。以恐龙作为标准来看,蜥脚下目是其中的巨无霸。在漫长的恐龙时代,即使是体型最小的蜥脚下目恐龙也要比它们栖息地内的其他动物要大,而最大的蜥脚类则比任何出现在地表的动物都要大出几个等级。
蜥脚类恐龙的体型巨大,所以其可能具有几项优势,例如防御掠食动物、减低能量的消耗、长寿,其中最重要的优势是消化食物。与小型动物相比,大型动物的消化作用的效率更高,因为食物在它们的消化系统中停留更多的时间。基于此,大型动物可用较低营养价值的食物维持生存。蜥脚类恐龙的化石,大部分发现于过去为干旱或季节性干旱的地层中,它们可以大量、低营养价值的植物为食,以在这些环境中维持生存。
在恐龙占优势的侏罗纪与白垩纪期间,几乎所有身长超过1米的陆地动物皆为恐龙,同时代的其他动物因此受到体型发展与生态位上的限制。举哺乳类为例,它们的体型多接近现代的啮齿动物,不超过猫的大小,食性为肉食性,以小型动物为生。生存在白垩纪的三椎齿兽目的巨型爬兽(Repenomamus giganticus)是个少数例外,巨型爬兽体重12到14公斤,已知会以幼年鹦鹉嘴龙等小型恐龙为食。
生理特征
自上个世纪60年代以来,恐龙的体温调控机制成为科学家们争论的主题之一。恐龙恒温动物说逐渐成主流理论,它们被视为活跃的动物,至少具有相当稳定的体温。目前的争论多在于它们的体温调节机制,以及它们与鸟类、哺乳类的代谢率相近程度。
早期对恐龙的研究中,古生物学家认为恐龙是类似蜥蜴的冷血动物。他们认为恐龙是群相当缓慢、慵懒的动物,需要外部热量以调节它们的体温,接近现代爬行动物。恐龙冷血动物说成为主流看法,直到1968年,罗伯特·巴克公布了一份革命性的研究,提出恐龙恒温动物说。
目前的证据多指出恐龙可生存在较寒冷的气候环境,且至少有某些恐龙借由内部身体机制来调节体温,例如巨大体型造成的巨温性。恐龙恒温动物说的证据包含:在南极洲与澳洲发现恐龙化石,两地区在当时位于极区内,有长达六个月的永夜、某些恐龙具有隔绝热量用的羽毛、以及骨头内具有恒温动物特有的心血管结构。从骨骼结构显示,兽脚类与其他恐龙是活跃的动物,若它们具有恒温动物的心血管系统,将符合这样的生活方式;而蜥脚类恐龙则拥有较少的恒温动物体征。最有可能的状况是,某些恐龙是恒温动物,而其他则不是。恐龙恒温/变温动物说的争论与新证据仍在持续进行中。
白垩纪灭绝事件
非鸟类恐龙消失于6550万年前的白垩纪-古近纪灭绝事件,同时间还有许多生物也跟着灭亡,包含:沧龙科、菊石、翼龙目、蛇颈龙目、植食性的乌龟与鳄鱼、大部分鸟类、以及原始哺乳类。自从1970年代以来,有许多研究试图探索这次灭绝事件的原因,并形成数个理论。通常的解释是一个撞击事件造成了灭绝,有些科学家提出其他理由,或者是几个综合事件导致这次灭亡。
在恐龙时代末期,极区并没有冰帽,海平面比现在高出100到250米。地球各地的气温差异不大,极区与赤道地区的气候相差不到25℃;整体而言较今日暖和,极区的气温高于现在的气温约50℃。
恐龙时代的大气成分也与今日有很大差别:二氧化碳是今日的12倍,氧气则占了大气的12%到18%,而今日则占了21%。但是在白垩纪晚期,地球的环境有显著的变动:火山活动增加,造成气候变冷。也有部分科学家进一步推测,气候变化是导致灭亡的原因之一。
小行星撞击事件
在1970年代后期,加州大学伯克利分校的路易斯·阿尔瓦雷茨提出了行星撞击理论,造成了白垩纪末期的大规模灭绝事件。阿尔瓦雷茨提出世界各地的地层,微量元素铱含量在6550万年前有大量的增加,可视为撞击的直接证据。有许多证据显示当时有颗直径5到15公里的陨石或彗星,撞击如今的犹加敦半岛地区,形成宽170公里的希克苏鲁伯陨石坑,引发此次灭绝事件。但科学家不确定恐龙是在撞击事件中灭亡,还是在此之前就已经衰退

化石纪录不能看出恐龙灭绝的速度,但许多不同的测试研究发现它们的灭亡相当迅速。这些结果使科学界更加相信行星撞击理论。撞击事件在短期内会造成高温,产生的扬尘会遮辟天空,造成全球性的长时间气候冷却。大部分植物因无法光合作用而失去功能,植食性恐龙也因没有食物而死亡,而肉食性恐龙也因没有食物来源,也渐渐灭亡,仅有小型动物和少量植物幸存下来。
在2007年9月,美国科罗拉多州美国西南研究院的研究人员威廉·波特克(William Bottke)与捷克籍的科学家,利用电脑计算出希克苏鲁伯撞击事件的可能原因。根据他们的动态模型表明,该陨石坑有90%的可能是由一个名为巴普提斯蒂娜的小行星所造成的,该小行星直径接近160公里,运行于火星与木星之间,火星和木星的引力会导致许多大物体被扰乱到穿越地球的轨道;在1亿6000万年前,巴普提斯蒂娜小行星被一颗直径约55公里的未命名小行星撞击后粉碎,主要的碎片形成巴普提斯蒂娜族小行星带,而某些碎片则闯入地球的公转轨道,与地球碰撞,其中一颗直径10公里的碎片,在6,550万年前撞击了墨西哥犹加敦半岛,形成希克苏鲁伯陨石坑,这些碎片可以解释白垩纪(从大约1.45亿年前到6500万年前)和随后的新生代时代的陨石坑率增加,这些碎片实际上会在内部太阳系中形成持续约一亿年的小行星阵雨。
威廉·波特克的研究结果与阿尔瓦雷茨的理论相似,另外也采纳了伯克利大学的物理学家查理·穆勒所提出的太阳伴星理论。天文学家查理·穆勒(Richard-A-Muller)提出太阳有颗名为复仇女神(Nemesis)的伴星,复仇女神每隔2600万年会经过奥尔特云彗星带,干扰了彗星的轨道,使众多彗星进入内太阳系。这些彗星的少部分会进入地球的公转轨道,与地球发生撞击,进而造成大规模的灭绝事件。
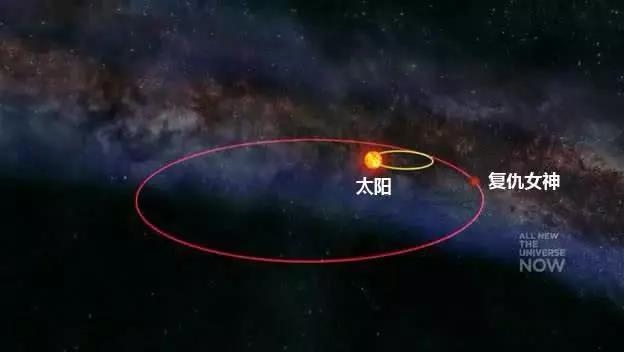
但科学家认为,事实上,地球生物大灭绝的规律性恰恰证明了“复仇女神”理论错误,因为与其它恒星的长期相互作用,“复仇女神”的轨道应该会发生改变。研究人员说:“曾经推动‘复仇女神’理论的化石数据推翻了该理论。”上一次大灭绝事件发生在1100万年前,它造成了地球上10%的生物灭绝。这意味着距离下次生物大灭绝事件还有1600万年,因为经研究发现地球生物每2700万年灭绝一次。经图表显示,有时灭绝事件最多会提前1000万年发生。
火山爆发
在2000年以前,就有学者主张德干暗色岩(Deccan Traps)是这次灭绝事件的主因,但因为这次火山爆发始于6,625万年前,并持续超过20万年,所以当时认为德干暗色岩火山爆发所造成的灭绝事件应该是逐步、缓慢的。但现在有证据显示,有2/3的德干暗色岩地形,是在一万年内形成的,发生于6,550万年前。显示这次火山爆发造成了快速的灭绝事件,可能在数千年内,但还是远比撞击事件长。
德干暗色岩火山爆发借由多种方式造成了灭绝事件,灰尘与二氧化硫大量喷出,灰尘使地表的日照量下降,植物的光合作用减少;当灰尘慢慢降落至地表时,火山爆发喷出的灰尘和二氧化硫气体,造成许多植物因此而死亡,植食性恐龙因没有食物而灭亡,而肉食性恐龙也相继灭绝。在恐龙大规模灭绝前,火山爆发喷出的气体,造成了气候变动,大气温度曾下降2°C。
在德干暗色岩火山爆发仍被认为是缓慢灭绝事件时,路易斯·阿尔瓦雷茨便已提出古生物学家被少数的资料所混淆。他的建议当时被没有被接受,但后来针对化石层的研究工作,证明早期的相关研究确实有误。目前大部分古生物学家已普遍接受撞击事件是白垩纪末灭绝事件的主要原因。不过,路易斯·阿尔瓦雷茨的儿子沃尔特·阿尔瓦雷茨认为在这次撞击事件前,地球上已发生其他重大事件,例如:海平面下降、造成德干暗色岩的大规模火山爆发,它们可能也是这次灭绝事件的综合原因之一。
科学家们发现,许多物种的灭绝是集中发生的,在很短的时间内,发生了大规模物种灭绝事件,且是存在周期性规律的,其周期约为2600万年。自寒武纪以来,在地球上已发生超过5次以上的生物大灭绝。尽管有人对这一数字表示质疑,但是,人们对这种周期性的灾难已经有了充分的认识。
1983年,物理学家对地球上的生物灭绝事件进行了研究。他们搜集了近2.5亿年以来的古生物资料,并用统计的方法进行了分析。研究结果表明,在这段时间里,地球上至少发生过7次大规模物种灭绝事件。其中,灭绝率最高的一次发生在白垩纪。在这次事件中,生物物种灭绝高达70%。正是发生在白垩纪的这场灾难,使得恐龙最终灭绝了。
搜狗科学:baike.sogou.com/kexue/d40527149689499136.htm
下一篇:碱基是什么?关于碱基的科普介绍