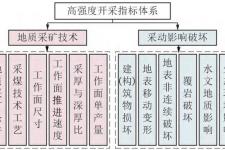
鹦鹉螺亚纲是什么?关于鹦鹉螺亚纲的科普介绍

鹦鹉螺类是海洋头足类(软体动物)的一大类,属于鹦鹉螺亚纲,始于晚寒武纪,今天以活着的鹦鹉螺和异鹦鹉螺为代表。鹦鹉螺类在古生代早期繁盛起来,那个时候它们构成了主要的食肉动物,并形成了异常多样的贝壳形状和形态。已知大约有2500种鹦鹉螺化石,但只有少数种类存活至今。
分类学关系
鹦鹉螺类是头足类动物中的一类,头足类是一种高级软体动物,也包括菊石、贝伦石和现代鞘类动物,如章鱼和鱿鱼。其他软体动物包括腹足类、舟状类和双壳类。
传统上,头足类最常见的分类是四重分类,分为直立角蟾、鹦鹉螺、菊石和鞘类。本文是关于广义上的鹦鹉螺亚纲的,有时称为Nautiloidea sensu lato。
从分支学角度来说,鹦鹉螺类是一种由衍生头足类中所没有的共有原始(准同形)特征结合在一起的复系集合。换句话说,它们是一个等级群,被认为产生了直角蟾、菊石和胚芽鞘,并通过排除这些后代群来定义。菊石和胚芽鞘在传统上都被认为是杆石类动物的后代,而杆石类动物又是直壳原角鲨烯(Orthoceratoids)的后代。
菊石(包括菊石和角闪石的一个群体)是鹦鹉螺已经灭绝了的近亲,早在大约4亿年前的泥盆纪就进化了。
一些工作者将鹦鹉螺亚纲这个名字应用于一个更专属的群体,严格意义上的鹦鹉螺亚纲。这个分类单元只包括那些与现代鹦鹉螺明显相关的目。分配的成员因作者不同而有所不同,但通常包括跗藻纲、盘尾丝虫和鹦鹉螺目。
特征
广义的鹦鹉螺亚纲有两个主要特征——简单的向前凹的隔膜,通常产生简单的缝线,以及隔膜颈部指向后部的吸管(即在动物的整个个体发育过程中是逆向虹吸的)。
辐锥腔(壳的腔室部分)之间的隔膜是在动物快速生长期间形成的。那时,套膜的后部分泌一种新的隔膜,增加了另一个腔室,而更靠前的部分增加了外壳。动物的身体,也就是它的内脏,继续占据外壳的最后一个腔室——住室。
隔膜被贯穿外壳每个内腔的吸管刺穿。在细管的肉质管周围是文石(碳酸钙的多晶型物——在石化过程中经常重结晶成方解石,一种更稳定的碳酸钙形式)构成的结构:隔膜颈部和连接环。一些早期鹦鹉螺类动物将碳酸钙沉积在空室(称为喀麦隆沉积物)或虹吸管(内胚层沉积物)中,这一过程可能与控制浮力有关。在对鹦鹉螺类进行分类时,虹吸管的性质及其在壳内的位置非常重要,有助于将其与菊石类区分开来(在大多数菊石类中,虹吸管位于壳的外围,而在大多数鹦鹉螺类中,它穿过腔的中心)。
缝线(或缝合线)在外壳表面上以一系列狭窄的波浪线可见,它们出现在每个隔膜接触外壳壁的地方。鹦鹉螺的缝线形状简单,要么直,要么稍微弯曲。这不同于棱菊石的“之字形”缝线和菊石的高度复杂缝线。
现代鹦鹉螺

关于已灭绝的鹦鹉螺的许多知识是基于我们对现代鹦鹉螺的了解,例如发现于从萨摩亚到菲律宾的西南太平洋和澳大利亚海岸外的印度洋的有腔鹦鹉螺。它通常不在100米以下的水中发现,可能在500到700米(1600到2300英尺)之下发现。
鹦鹉螺是自由游动的动物,有一个头,有两只简单的无晶状体眼睛和手臂(或触须)。它们有一个光滑的外壳覆盖在一个大的身体腔室上,身体腔室被分成充满惰性气体的子腔室(类似于大气的成分,但是氮气更多,氧气更少),使动物在水中呈中性浮力。多达90根触须围绕口腔排成两个圆圈。这种动物是食肉动物,有角质和喙状的下颚,允许它以甲壳类动物为食。
空鹦鹉螺壳可能会漂很远,日本、印度和非洲都有报道。毫无疑问,这同样适用于鹦鹉螺化石的外壳,在动物死后,外壳内的气体使其在一段时间内保持浮力,允许空外壳在最终沉入海底之前从动物居住的地方被带出一段距离。
鹦鹉螺通过喷射力来推进自己,从一个叫做海波门的细长漏斗中排出水,这个漏斗可以指向不同的方向来控制它们的运动。与箭石类和其他头足类动物不同,现代鹦鹉螺没有墨囊,也没有证据表明灭绝的物种也有墨囊。此外,与已灭绝的菊石不同,现代鹦鹉螺没有口盖,也没有任何种类的板来封闭壳。除了一个例外,在任何灭绝的鹦鹉螺身上也没有发现这样的板块。
现代鹦鹉螺壳的颜色非常突出,虽然有些罕见,但已知壳的颜色保存在鹦鹉螺化石中。它们通常只在背侧显示颜色图案,这表明活着的动物是水平游动的。
化石记录



鹦鹉螺类通常在古生代早期岩石中作为化石被发现(在更近的地层中就不那么常见了)。鹦鹉螺化石的壳可以是直的(即直锥的,如直角石和瑞诺角石),弯曲的(如弓角石),卷曲的(如新生角石),也很少是螺旋的(如洛里角石)。一些物种的壳——特别是在古生代晚期和中生代早期——用刺和肋骨装饰,但大多数都有光滑的壳。
贝壳由文石形成,尽管喀麦隆沉积物可能由原生方解石组成。
波罗的海沿岸和美国部分地区奥陶系时期的岩石含有多种鹦鹉螺化石,在爱尔兰石炭纪的石灰岩中可能发现了圆盘角石(Discitoceras)和瑞诺角石(Rayonnoceras)等标本。英国侏罗纪时期的海相岩石经常产生新生角石的标本,在美国中北部白垩纪的皮埃尔页岩(Pierre Shale)地层中也发现了外扩角石等鹦鹉螺类。
奥陶纪鹦鹉螺类内角石的标本已经被记录下来,最长可达3.5米(11英尺),而房角石类(有点怀疑)估计可达11米(36英尺)。这些大型鹦鹉螺类动物在它们活着的时候一定是其他海洋动物的可怕掠食者。
在一些地方,如斯堪的纳维亚和摩洛哥,正圆锥形鹦鹉螺化石堆积如此之多,以至于它们形成了由非特异性化石组成的石灰岩,在地质文献中称为头足类地层、头足类石灰岩、鹦鹉螺石灰岩或正圆锥形石灰岩。虽然直角石属这个术语现在只指波罗的海沿岸奥陶系的一个属,但在以前,它被用作生活在奥陶系到三叠纪(但在早古生代最常见)的所有直壳鹦鹉螺的总称。
进化史
鹦鹉螺类最早发现在中国东北凤山县,该化石是寒武纪晚期形成的,在那里它们似乎非常多样(当时这是一个温暖的浅海,富含海洋生物)。然而,尽管已经从131个命名的物种中提出了四个目,但并不确定所有这些都是有根据的,事实上这些分类群很可能严重过度分类了。

这些早期种类中的大部分都灭绝了,但是一个单独的家族——爱丽斯曼角石科——存活到了奥陶纪早期,在那里它最终产生了所有后来的头足类动物。奥陶纪早期和中期鹦鹉螺经历了辐射演化。 此时出现了大约八个新的目,涵盖了种类繁多的贝壳类型和结构,以及生态生活方式。
鹦鹉螺类在奥陶纪、志留纪和泥盆纪一直保持着较大的适应范围和形态多样性,各种直的、弯曲的和盘绕的壳形态同时并存。在那段时间里,早期的几个目灭绝了,但其他的目变得繁荣起来。
鹦鹉螺类在泥盆纪开始衰落,可能是由于与它们的后代和亲属——菊石类和鞘类——的竞争,只有鹦鹉螺类拥有自己的物种(事实上,多样性在增加)。它们的壳变得越来越紧密盘绕,而非鹦鹉螺物种的数量和种类在整个石炭纪和二叠纪都在持续减少。
二叠纪末期的大规模灭绝对鹦鹉螺类的损害小于对其他类群的损害,少数类群存活到中生代早期,包括伪角类、杆石类、鹦鹉螺类和可能的直角石类。人们长期以来认为最后的直壳形式在三叠纪末消失了,但是在白垩纪岩石中发现了一种可能的正长石类。除了这个例外,只有一个鹦鹉螺亚目,延续到整个中生代,在那里它们与更特别的菊石类表亲非常愉快地共存。这些形式中的大多数与现代鹦鹉螺略有不同。它们在第三纪早期曾短暂复苏(也许填补了白垩纪末期菊石灭绝时留下的生态位),并一直保持着全球分布,直到新生代中期。随着中新世和上新世全球变冷,它们的地理分布缩小,这些耐寒长寿的动物的多样性再次下降。今天只有六种活着的物种,都属于鹦鹉螺(珍珠鹦鹉螺)和异鹦鹉螺两个属。
分类
分类各不相同,并且随着新信息的发现和不同工作者的观点而变化。泰瑟-1988年的分类是另一个分类,泰瑟等人1964年在《论文第K部分》中的分类也是另一个分类。
韦德(1988)将鹦鹉螺亚纲分为6个超目,结合了系统发育相关的目。它们是:
- 瓜角石亚纲=短棒角石目、原珠角石目、沿河角石目和丽斯木角石目。
- 内角石亚纲 内角石目直角石亚纲=直角石目、袋角石目和假直角石目(krger 1007的直角石目)
- 鹦鹉螺亚纲=触环角石目、箭钩角石目和鹦鹉螺目。
- 珠角石亚纲=珠角石目
- 迪斯科亚纲 迪斯科目
它们中的三个被建立为分类内角石目、珠角石目和迪斯科目的相同位置。共享一个共同祖先并形成鹦鹉螺分类树一个分支的三个联合相关的目;瓜角石亚纲,主要由小寒武纪组成,包括后来种群的祖先;直角石亚纲,它结合了不同的主要直圆锥目,其中一个是杆石目和菊石的来源;鹦鹉螺纲,包括第一批卷形头足类动物——触环角石目,以及鹦鹉螺目,包括现在的鹦鹉螺。
另一个目,杆石目(Bactritida),来源于直角石亚纲,有时包含在鹦鹉螺亚纲中,有时包含在菊石亚纲中,有时归入它自己的亚类,杆石亚纲。
最近,该领域的一些工作者已经认识到迪斯科目是一个独特的序列,还有假直角石目,两者以前都被列为直角石目的子序列。
另一种解释(恩格尔,1997-1998年)认为鹦鹉螺类,实际上是头足类,分为两大类,帕尔头足类(包括除直角石亚目和袋角石目以外的所有鹦鹉螺类)和新头足类(头足类的其余部分)。然而,林德格伦等人(2004年)最近的系统发育研究支持头足类的单系性,鹦鹉螺亚纲是箭鞘亚纲的姐妹群。
搜狗科学:baike.sogou.com/kexue/d10047.htm