遗传学是什么?关于遗传学的科普介绍

遗传学是生物学的一个分支,研究生物体中的基因、遗传变异和遗传
虽然遗传已经被观察了几千年,但是直到19世纪,一位叫孟德尔的科学家、奥古斯丁 修士,对遗传学首次进行了科学研究。孟德尔研究的“性状遗传”是一种性状从父母传给后代的方式。他观察到生物体(豌豆植物)通过独立的的“遗传单位”将性状进行遗传。遗传学中至今仍在使用遗传单位”这个术语,它也是一个有点模糊的基因定义。
基因的性状遗传和分子遗传机制仍然是21世纪遗传学的基本原理,但现代遗传学已经不再仅仅是对遗传进行研究而扩展到对基因的功能和行为进行研究。基因结构和功能、变异和分布是在细胞、生物体(例如种群)和群体的背景下开展研究的。遗传学分为很多领域,例如表观遗传学和群体遗传学。遗传学所研究的生物体遍布整个生命领域(古细菌、细菌和真核生物)。
遗传过程与生物体所处的环境和经历相结合,共同对发育和行为产生影响,通常被称为先天与后天。细胞或生物体的细胞内或细胞外环境可以起始或关闭基因的转录过程。经典的例子是两种基因相同的玉米种子,将一种放置在温带气候中,另一种放置在干旱气候中。虽然在遗传上两个玉米秸秆的应表现出相等的平均高度,但在干旱气候中的玉米秸秆由于生长环境中缺乏水和营养,只能达到在温带气候中生长的玉米秸秆一半的高度。
语源
遗传学源于古希腊 γενετικός genetikos意思是“属格”/“生成”,这反过来又来源于γένεσις genesis,意思是“起源”。
历史
从史前时代起,生物从父母那里继承性状的现象就被用来进行选择性育种来对作物植物和动物进行改良。对这一过程进行科学理解的现代遗传学是在19世纪中叶由一名奥古斯丁的工作 修士 的孟德尔开创的。
在孟德尔之前, imre festics一名住在Kőszeg的匈牙利贵族,他是第一个使用“遗传学”这个词的人,他在作品《自然的遗传法则》(《自然遗传》,1819年)中描述了几个有关基因遗传的规则。他的第二定律与孟德尔发表的观点相同。在他的第三定律中,他对突变的基本原理进行了拓展(他可以被认为是雨果·德·弗里斯的先驱)。

孟德尔的工作之前还有其他遗传理论。在19世纪流行的一个理论,由查尔斯·达尔文在1859提出的物种起源,即融合遗传:认为个人从父母那里继承了一系列特征。孟德尔的工作提供了杂交后性状绝对不会融合的例子,表明性状是由不同基因组合形成的,而不是由连续的混合所产生的。后代性状的混合现在可以解释为多个基因产生的定量效应。当时另一个广泛认可的理论是后天特征的遗传:个人继承父母所具有的性状。这个理论(通常与让-巴蒂斯特·拉马克联系在一起)现在被认为是错误的——个人的经历不会对传递给孩子的基因产生影响,尽管在表观遗传学领域的一些证据验证了拉马克理论的某些方面的合理性。其他理论包括查尔斯·达尔文的《泛生》(既有些性状即是先天的也是后天的)和弗朗西斯·高尔顿对泛生重新定义为既有颗粒性又有遗传性的表述。
孟德尔和经典遗传学
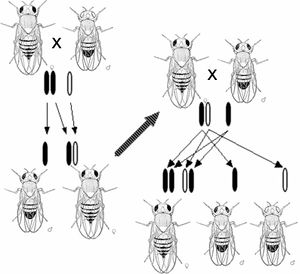
现代遗传学始于孟德尔对植物的遗传性质进行研究。在他于1865年提交给Naturforschender Verein(自然研究学会)的论文”versucheüber Pfranzenybriden“(“植物杂交实验”)中孟德尔描述了豌豆植株某些性状的遗传模式,并对其进行了数学描述。虽然这种遗传模式只能观察到少数性状,但孟德尔的工作证明遗传是微粒状的,而不是由后天产生的,许多性状的遗传模式可以通过简单的规则和比率来进行解释。
孟德尔的工作直到1900年雨果·德·弗里斯和其他科学家重新对他的发现进行了研究,才使得孟德尔的理论受到了广泛的关注。孟德尔研究的支持者威廉·贝特森在1905年创造了遗传学这个词(形容词,意思为起源的,源自希腊语单词发生—γένεσις,“起源”在这个名词出现前就已经在使用了,并于1860年首次在生物学意义上使用)。巴特森既是这一领域的先驱者,同时他也得到剑桥纽纳姆学院其他科学家的大量帮助,特别是贝基·桑德斯、诺拉·达尔文·巴罗和穆里尔·惠尔代尔·翁斯洛。巴特森普在1906年于伦敦举行的第三届国际植物杂交大会上的开幕词中推广了遗传学这个词用于描述遗传研究的用法。
孟德尔的工作被重新发现后,科学家们试图确定出细胞中的哪些分子负责遗传。1911年,托马斯·亨特·摩尔根基于对果蝇性别相关的白眼突变的观察,得出结论认为基因位于染色体上。1913年,他的学生阿尔弗雷德·斯特蒂文特利用遗传连锁现象表明基因在染色体上呈线性排列。
分子遗传学
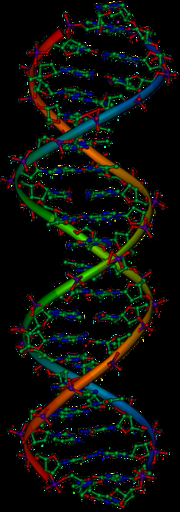
虽然已经确定基因位于染色体上,但染色体由蛋白质和DNA组成,科学家仍不能确定这两者中哪一种物质负责信息的遗传。1928年,弗雷德里克·格里菲斯发现了转化现象(见格里菲斯实验):死细菌可以通过转移遗传物质来“转化”其他活细菌。1944年,埃弗里-麦克劳德-麦卡蒂实验确定DNA就是负责转化的分子。1943年,哈默林在对单细胞藻类伞藻属的研究中确认细胞核是真核生物遗传信息储存库。1952年的赫雪-蔡司实验证明了DNA(而不是蛋白质)是感染细菌的病毒的遗传物质,这也进一步证明了DNA是负责遗传的分子。
1953年詹姆斯·杜威·沃森和弗朗西斯·克里克年利用罗莎琳·富兰克林和莫里斯·威尔金斯的 X射线晶体学工作成果确定了DNA的结构,提出DNA具有螺旋状结构(即形状像开瓶器)。他们的双螺旋模型有两条核苷酸指向内部的DNA链,每条链与另一条链上的互补核苷酸相匹配,形成看起来像扭曲梯子上的梯级的结构。这种结构表明遗传信息存在于每条DNA链上的核苷酸序列中。该结构还提出了一种简单的复制方法:如果两条链发生分离,每条链都可以基于旧链的序列来合成新的伴侣链。这一特性赋予了DNA半保守性,即一条新DNA链来自原始父链。
尽管DNA的结构表明了遗传是如何进行的,但是仍不清楚DNA是如何对细胞的行为产生影响的。在接下来的几年里,科学家们试图理解DNA是如何对蛋白质生产产生控制的。人们发现细胞以DNA作为模板来合成对应的信使RNA ,信使RNA分子中的核苷酸序列与DNA非常相似。信使RNA中的核苷酸序列用于在蛋白质中创建一个氨基酸序列;核苷酸序列和氨基酸序列之间的转换被称为遗传密码。
随着对遗传的新的分子层面理解的加深,研究出现了爆炸式的发展。在1973年太田朋子提出了一个著名的理论,她对分子进化的中立原理进行修改发表了分子进化的近中立原理。在这个理论中,Ohta强调了自然选择和环境对遗传进化速度的重要性。一个重要的发展是1977年由弗雷德里克·桑格进行的链终止脱氧核糖核酸测序。这项技术使得科学家能够对DNA分子的核苷酸序列进行读取。1983年,凯利·班克斯·穆利斯开发了聚合酶链式反应,提供了一种快速从混合物中分离和扩增特定DNA片段的方法。人类基因组计划、美国国家卫生研究院能源部的努力和赛雷拉基因公司 的努力使得在2003年人类基因组的测序的成果。
遗传的特征
离散遗传和孟德尔定律
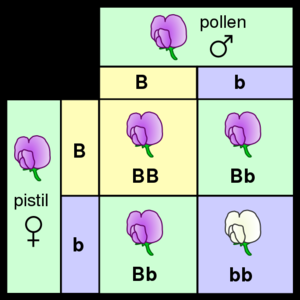
本质上来说,生物遗传是通过将称为基因的离散可遗传单位从父母传给后代来实现的。孟德尔通过研究豌豆植物遗传性状的分离,发现出了这一特性。孟德尔在对花颜色特征进行探究的实验中观察到,每种豌豆的花要么是紫色的,要么是白色的,但是从来没有出现两种颜色之间的颜色。同一基因的不同的离散被称为等位基因。
以豌豆为例,豌豆是一种二倍体,每株植物的基因都有两个拷贝,其中一个从亲本遗传得来的。包括人类在内的许多物种都根据这种遗传模式进行遗传。具有一个给定基因的相同等位基因的二倍体生物在该基因位点被称为纯合生物,而给定基因的两个等位基因不相同的生物被称为杂合生物。
某一特定生物的等位基因被称为基因型,而生物体的可被观察到的性状被称为其表现型。当生物体的某一个基因为杂合时,通常其中一个等位基因被称为显性,因为它可以决定生物体的表型,而另一个等位基因被称为隐性,因为它的性质退化并且不会被观察到。某些等位基因不具备完全显性,而是表现出一种中间表型因而具有不完全显性,或者通过同时表达两个等位基而具有共显性。
当一对生物体进行有性繁殖时,它们的后代会随机的从父母那里随机个继承继承两个等位基因中的某一个。这些对离散遗传和等位基因分离的观察结果统称为孟德尔第一定律或分离定律定律。
符号和示意图
遗传学家使用示意图和符号来对遗传进行描述。用一个或几个字母代表某个基因。通常用“+”来标记常见非突变等位基因。
在受精和育种实验中(特别是在讨论孟德尔定律时),父母被称为“P”代,后代被称为“F1”(子一代)。当F1代自交时,得到的后代被称为“F2”(子二代)。预测杂交育种结果通常使用旁氏表。
对人类遗传病进行研究时,遗传学家通常使用系谱图来表示性状的遗传。这些图表描述了一个家族树中某个性状的遗传情况。
多基因相互作用
生物体中含有数千个基因,在通过有性生殖进行繁衍的生物体中,这些基因通常相互独立分类。这意味着黄色或绿色豌豆颜色的等位基因的遗传与白色或紫色花的等位基因的遗传是不相关的。这种现象被称为“孟德尔第二定律”或“独立分配定律”,意味着不同基因的等位基因在父本与母本之间进行混合,从而形成具有多种不同组合的后代。(一些基因并不独立地进行排列,从而显示出遗传连锁,这是接下来需要讨论的主题。)
通常不同基因之间会以影响相同性状的方式产生作用。在蓝眼睛玛丽(琉璃草属·维纳例如,存在一个决定花颜色的等位基因:蓝色或洋红色。然而,另一个基因可以控制花朵到底是含有颜色还是白色。当一个植物含有两个白色等位基因时,它的花是白色的—无论第一种基因决定出现的颜色是蓝色还是品红色。这种基因之间的相互作用被称为上位性,这里第二个基因上位于第一个基因。
然而有许多特征不是离散(例如紫色或白色花朵),而是连续的(例如人类身高和肤色)。这些复杂性状是多种基因共同作用的结果。生物体所处的环境可以对这些基因产生一定影响。生物体的基因对复杂性状的影响程度被称为遗传力。性状遗传力的测量是相对的—在一个可变性较大的环境中,环境对性状的总变异有较大影响。例如,人的身高是一个由多种因素共同决定的复杂性状。比如在美国有89%的遗传力。然而,在尼日利亚,人们获得良好营养和保健的方法不同,身高的遗传力只有62%。
遗传的分子基础
DNA和染色体
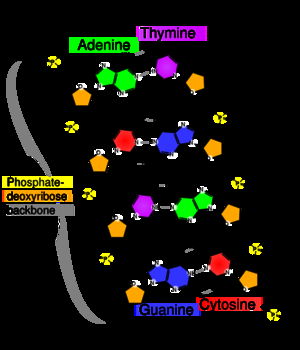
基因的分子基础是脱氧核糖核酸(DNA)。脱氧核糖核酸由核苷酸链组成,其中包括四种不同类型:腺嘌呤(A)、胞嘧啶(C),鸟嘌呤(G)和胸腺嘧啶(T)。遗传信息存在于这些核苷酸的序列中。 病毒是唯一不符合这一规则的物质—有些病毒可以使用RNA 而不是DNA作为它们的遗传物质。病毒在没有宿主的情况下无法进行繁殖,并且不受多种遗传过程的影响,因此往往认为不是活的生物体。
DNA分子通常以双链的形式存在,卷曲成双螺旋的形状。DNA中的每个核苷酸优先与相对链上的互补核苷酸进行配对,遵循的配对规则是:A与T配对,C与g配对。因此,在双链结果中,每个链都可以包含所有必要的信息,因此互补链的信息是多余的。DNA的这种结构是遗传的物理基础: DNA复制是通过将双链分裂,并分别使用每条链作为模板合成新互补链来复制遗传信息。
基因沿着DNA碱基对序列的长链呈线性排列。在细菌,每个细胞通常含有一个圆形基因带,而真核生物体(如植物和动物)的DNA通常排列成线性。通常这些DNA链非常长;例如,人类最大的染色体约有2 . 47亿个碱基对。 染色体的DNA与结构蛋白相关联,结构蛋白组织、压缩和控制对DNA的访问,形成一种叫做染色质的物质;在真核生物中,染色质通常由核小体组成,核小体是缠绕在组蛋白核周围的DNA片段。生物体内的全部遗传物质(通常是所有染色体的DNA组合序列)被称为基因组。
单倍体生物体每个染色体只有一个拷贝,大多数动物和植物都是二倍体生物,即每条染色体都是成对存在的,因此每个基因包含两个拷贝。一个基因的两个等位基因位于两个同源染色体的相同位点,每个等位基因从不同的亲本遗传获得。
许多物种都含有性染色体,来决定生物体的性别。在人类和许多其他动物中, Y染色体包含触发特定雄性特征发育的基因。在进化过程中,这条染色体失去了大部分信息和大部分基因,而 X染色体与其他常染色体相似,含有许多基因。X和Y染色体形成高度异质性的染色体对。
生殖
当细胞分裂时,首先复制整个基因组,每个子细胞获得其中的一个拷贝。这个分裂过程叫做有丝分裂,是最简单的生殖形式,也是无性生殖的基础。无性生殖也可以在多细胞生物中发生,从而产生从单亲那里遗传基因组的后代。与父母基因相同的后代被称为克隆。
真核生物通常使用有性生殖来产生包含有来自两个亲本混合遗传物质的后代。有性生殖的过程在单个基因组拷贝(单倍体)和双拷贝(二倍体)两种形式之间交替进行。单倍体细胞融合并结合遗传物质,形成具有成对染色体的二倍体细胞。二倍体生物通过只进行分裂,而不进行DNA复制的方式来形成单倍体,从而产生随机继承每对染色体中一个染色单体的子细胞。大多数动物和植物在其大部分生命周期中都是二倍体,单倍体形式存在于例如精子或者卵子这样的单细胞配子中。
虽然细菌不能使用单倍体/二倍体的方法进行有性生殖,但它们有许多获得新遗传信息的方法。一些细菌可以通过接合,即将一小块质粒DNA转移到另一种细菌中。 细菌也可以吸收存在于环境中的原始DNA片段,并将其整合到自己的基因组中,这种现象被称为转化。 这些过程导致基因水平转移,即在原本相互独立的生物体之间传递遗传信息。
重组和遗传连锁
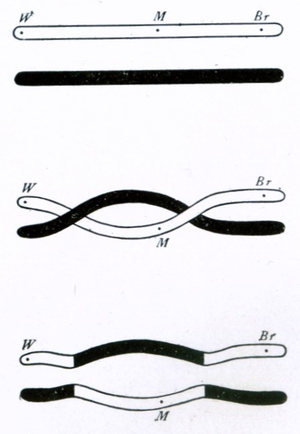
染色体的二倍体性质使位于不同染色体上的基因在形成单倍体配子的有性生殖过程中是相互独立或与其同源染色体对之间进行分离。这样,在交配对的后代中可以出现新的基因组合。理论上来说位于同一条染色体上的基因永远不会发生重组。然而它们可以通过染色体互换这一过程发生重组。在互换过程中,染色体交换DNA片段,有效地重组了染色体之间的基因等位基因。 染色体互换通常发生在减数分裂过程中,通过一系列的细胞分裂产生单倍体细胞。
1931年哈里特·克雷顿和芭芭拉·麦克林托克首次对染色体互换开展了细胞学研究。他们在玉米上进行的研究为遗传学理论研究提供了细胞学证据,该理论认为成对的染色体上的连接基因实际上会从一个同源染色体的一个位置交换到另一个同源染色体上。
染色体上两个给定点之间发生染色体互换的概率与点与点之间的距离有关。对于距离足够长的基因,交叉的概率足够高以至于使基因在事实遗传中是不相关的。然而,对于距离较近的基因,发生交叉的概率较低,这意味着遗传连锁的出现;这两个基因的等位基因往往是一起进行遗传的。一系列基因之间的连锁可以组合成一个线性的连锁图,该图谱大致描述了基因在染色体上的排列。
基因表达
遗传密码

基因通过产生蛋白质来表达表达它们的功能,蛋白质是负责细胞中大多数功能的复杂分子。蛋白质由一条或多条多肽链组成,每条多肽链由氨基酸根据一定的序列排列组成,根据基因的DNA序列(通过RNA中间体)被用来产生特定的氨基酸序列。这个过程开始于核糖核酸分子的序列与基因的DNA序列进行匹配,这个过程称为转录。
然后,该信使RNA分子通过翻译过程产生相应的氨基酸序列。序列中的每一组包含三个核苷酸,被称为密码子,每个密码子对应于蛋白质中包含的20种可能的氨基酸中的一种或终止子;这写密码子被称为遗传密码。这种信息的传递是单向的:信息从核苷酸序列转移到蛋白质的氨基酸序列中,但它不能将信息从蛋白质传递到DNA序列——这种现象在被弗朗西斯·克里克称为中心法则 。
氨基酸的特定序列形成蛋白质特定的三维结构,而蛋白质特定的三维结构与蛋白质的功能相关。有些蛋白质是简单的结构分子,如胶原蛋白形成的纤维。蛋白质还可以与其他蛋白质或小分子物质结合,有时可以通过促进结合分子内(不改变蛋白质本身的结构)的化学反应而起到类似酶的作用。蛋白质结构是可以发生改变的;血红蛋白通过弯曲成略微不同的形式,有利于在哺乳动物血液中捕获、运输和释放氧分子。
DNA中的单核苷酸差异可以导致蛋白质氨基酸序列发生改变。蛋白质结构是其包含的氨基酸序列产生的结果,可以通过破坏蛋白质结构的稳定或改变蛋白质表面以改变其与其他蛋白质和分子的相互作用来使蛋白质的性质发生改变。例如,人类一种遗传病,镰状细胞性贫血是由血红蛋白β-珠蛋白部分编码区内的单个碱基产生差异而导致一个氨基酸的类型发生改变,从而使血红蛋白的物理性质发生改变。镰刀型血红蛋白可以相互粘连,堆积形成纤维,导致携带镰刀型血红蛋白质的红细胞的形状发生改变。这些镰刀形的细胞不能流畅地通过血管进行运输,可能会堵塞血管或发生降解,最终导致某些相关医学问题的出现。
某些DNA序列虽然被转录成RNA,但没有被翻译成蛋白质——这种RNA分子被称为非编码RNA 。在某些情况下,这些物质会折叠生成参与关键细胞功能的结构(例如核糖体核糖核酸和转移核糖核酸)。RNA也可以通过与其他RNA分子(例如,microRNA )的相互作用来发挥调节作用。
遗传与环境

虽然基因包含生物体所以用于功能的信息,但环境在决定生物体最终的表型中也起着不可或缺的作用。短语“先天与后天”指的就是这种互补关系。生物体的表型取决于基因和环境的相互作用。比如暹罗猫皮毛的颜色。猫的体温对毛发的颜色起到重要作用。猫的基因编码产生深色的毛,因此猫的毛发生成细胞产生细胞蛋白质,产生深色毛发。但是这些产生深色毛发的蛋白质对温度十分敏感(即会发生由温度导致的突变),并且在高温环境中容易发生变性,在猫体温较高的区域不能产生深色毛发色素。然而在低温环境,蛋白质的结构是稳定的,通常会产生深色毛发的色素。这种蛋白质在较冷的皮肤区域可以保持功能—例如它的腿、耳朵、尾巴和脸—所以猫的四肢有深色的毛发。
环境因素对人类遗传病苯丙酮尿症起着重要影响。导致苯丙酮尿症出现的突变会破坏身体分解氨基酸苯丙氨酸的能力,导致中间分子的积聚,进而导致出现严重的智力障碍和癫痫症状。然而,如果患有苯丙酮尿症的患者严格控制饮食,避免摄入这种氨基酸,他们就会保持健康的身体状态。
确定基因和环境(“先天和后天”)对表型影响的常见方法包括研究同卵和异卵双胞胎,或多胞胎的其他兄弟姐妹。同卵兄妹的基因是相同的,因为他们来自同一个受精卵。同时,异卵双胞胎和正常的兄弟姐妹在基因上是不同的。通过比较某一疾病在同卵双胞胎中发生的频率和在异卵双胞胎中发生的频率,科学家可以确定该疾病是由遗传因素导致的还是由后天的环境因素引起的。一个著名的例子涉及对 基因四胞胎的研究,他们是同卵四胞胎,并都被诊断患有精神分裂症。然而,这种测试不能将影响胎儿发育的遗传因素与环境因素分开。
基因调节
通常生物体的基因组包含数千个基因,但这些基因不需要时刻保存活跃。当基因被转录成mRNA时进行表达并且细胞中存在许多控制基因表达的方法,只细胞需要时才产生相应蛋白。转录因子是与DNA结合的调节蛋白,可以促进或抑制基因的转录。在大肠杆菌的基因组中,存在一系列合成色氨酸所必需的基因。然而,当细胞中存在色氨酸时,这些色氨酸合成基因就处于沉默状态。色氨酸的存在直接对基因的活性产生影响——色氨酸分子与色氨酸阻遏物(一种转录因子)结合,改变阻遏物的结构,使得阻遏物与基因结合。色氨酸阻遏物阻断基因的转录和表达,从而对色氨酸的合成过程产生负反馈调节。

基因表达的差异在多细胞生物中尤其明显,因为在多细胞生物中,每个细胞都含有相同的基因组,但由于基因组表达的不同,它们具有不同的结构和行为。多细胞生物中的细胞都是由单个细胞产生的,根据所处环境和细胞间信号的不同分化成不同的细胞类型,并逐渐建立不同的基因表达模式,从而产生不同的行为。由于没有单个基因负责控制多细胞生物体内结构的发育,这些模式源于许多细胞之间的相互作用。
在真核生物中,存在影响基因转录的染色质结构特征,通常以DNA和染色质修饰的形式被子细胞稳定遗传。这些特征被称为”表观遗传,因为它们存在于DNA序列的“顶部”,并在下一代细胞中继续遗传。由于表观遗传的特征,在同一培养基中生长的不同类型的细胞可以保留不同的特性。虽然表观遗传特征在发育过程中通常是动态的,但有些特征如副突变,可以多代遗传,并且作为DNA作为遗传基础的生物中一般规则以外的罕见规则而存在。
遗传改变
突变
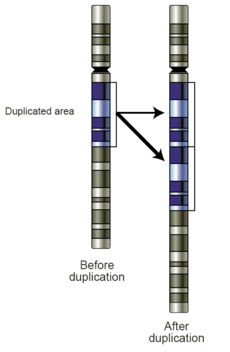
在DNA复制过程中,第二条链的合成可能会发生错误。这些被称为突变的错误可能会对生物体的表型产生影响,尤其是当它们发生在基因的蛋白质编码区中。由于 DNA聚合酶的“校对”功能错误率通常非常低——每一千万到一亿个碱基对可能会出现一个错误。 增加DNA突变几率的过程被称为诱变:化学品可以通过干扰碱基配对结构来特高DNA复制中的错误率,而紫外线通过破坏DNA结构来诱导突变的发生。 DNA中也可能出现自然的化学损伤,细胞中的 DNA修复机制可以修复产生的错配和断裂。然而,修复并能完全恢复原始序列。
在发生染色体互换DNA和重组基因的生物体中,减数分裂过程中的排列错误也可能导致突变的发生。 当相似的序列导致配对染色体采用错误配对时,错误很可能发生交叉互换中;这使得基因组中的一些区域更可能以这种形式发生突变。这些错误造成了DNA序列发生巨大的结构变化 – 整个区域的复制、倒置、缺失 – 或者不同染色体之间整个序列的错误交换(染色体易位)。
自然选择和进化
突变可能会对生物体的基因型产生改变,也可能会导致表型发生改变。大多数突变对生物体的表型、健康或生殖适度不会产生太大影响。突变通常是有害的,但有少量可能是有益的突变。对果蝇的研究表明,如果一个突变使基因产生的蛋白质发生改变,大约70%的突变是有害的,其余的突变要么是中性的,要么是微弱的有益的。
群体遗传学研究群体内遗传差异的分布以及这些分布如何随时间变化。 群体中等位基因频率的变化主要受自然选择的影响,其中等位基因为生物体提供选择性或生殖优势, 以及其他因素,例如突变、遗传漂变、遗传搭车, 人工选择和迁移。
经过许多代,生物体的基因组可以发生显著变化,从而导致进化的发生。在适应过程中,有益突变的积累可以使物种进化成在环境中更适于生存的形式。 新物种是通过物种形成产生的,这通常是由于地理上的分离阻止了种群之间的基因交流。
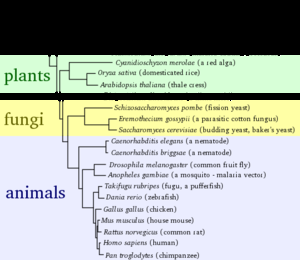
通过比较不同物种的基因组之间的同源性,可以计算它们之间的进化距离以及它们何时可能发生了分化。通常认为遗传是用来描述物种间亲缘关系的一种比表型特征比较更为准确的方法。物种之间的进化距离可以用来进化树来表示。这些树代表了共同起源和物种随时间的分化,虽然它们没有显示不相关物种之间遗传物质的转移(称为基因水平转移,在细菌中较为常见)。
模式生物

最初遗传学家会对各种生物体的遗传进行研究,但研究人员开始针对性的研究特定生物体某一子集的遗传。对某种生物体进行大量的先期研究,将鼓励新的研究人员在它的基础上进行更深层的研究,因此模式生物成为大多数遗传学研究的基础。模式生物中开展的研究课题包括基因调控的研究以及基因在发育和癌症发生中的参与。
选择这样的模式生物进行研究—部分原因是它们的繁殖时间短,而且易于基因操作,这使得这样的生物体成为进行遗传学研究工具。广泛使用的模式生物包括大肠杆菌,拟南芥,面包酵母(酿酒酵母),秀丽隐杆线虫,普通果蝇(果蝇),还有普通家鼠(肌肉鼠)。
医学
医学遗传学试图发现遗传变异与人类健康和疾病之间的关系。当寻找可能与疾病相关的未知基因时,研究人员通常使用遗传连锁和遗传谱系图来寻找与疾病相关的基因组位置。在群体水平上,研究人员利用孟德尔随机法在基因组中寻找与疾病相关的位置,这种方法对研究单个基因没有明确定义的多基因性状特别有用。一旦找到候选基因,通常对模型生物的相应(或同源)基因进行进一步的研究。除了研究遗传疾病外,基因分型方法的增加还导致了药物遗传学领域的出现:研究基因型是如何影响药物反应的科学。
个体发展成癌症的遗传倾向不同,癌症是一种遗传疾病。体内癌症发展的过程是由多种因素共同影响的。当细胞分裂时,可能会在体内细胞内发生突变。虽然这些突变不会遗传给后代,但它们可以影响细胞的行为,有时会导致细胞更频繁地生长和分裂。生物体内存在的一些机制会阻止这一过程的发生;向分裂失控的细胞传递特定信息,诱导细胞死亡,但有时会发生其他突变,导致细胞对这些信息敏感。在体内发生自然选择,最终在细胞内积累这些突变,促进细胞生长,形成恶性肿瘤并侵入身体各种组织。
正常情况下一个细胞只根据生长因子提供的信息进行分类,一旦与周围细胞接触就停止生长并且接收到生长抑制信号时细胞也会停止生长。通常细胞的分裂次数是有限的并且会死亡,始终停留在上皮细胞控制的范围内,不会迁移到其他器官。要癌变形成癌细胞,细胞必须在基因中积累突变(三到七个)。癌细胞可以在没有生长因子的情况下进行分裂,并不受抑制信号的控制。此外癌细胞具有不死性,甚至在与相邻细胞接触后也可以持续生长。癌细胞可以从上皮细胞限制的范围内脱离,最终从原发瘤发生转移。然后发生转移的癌细胞可以穿过血管内皮,被血流运送到新的器官,从而形成转移。虽然在一小部分癌症中具有一定的遗传倾向,主要是由于出现的一组新的遗传突变,这些突变最初出现在一个或少数细胞中并逐渐积累,最终形成肿瘤,但是这种突变并不遗传给后代(体细胞突变)。最常见的突变是肿瘤抑制基因p53蛋白或p53信号通路功能丧失,以及Ras蛋白,或其他致癌基因的功能发生突变。
研究方法
可以在实验室中对DNA进行操作。限制性内切酶是一种常用的酶,它在特定序列上切割DNA,产生可预测的DNA片段。可以通过使用凝胶电泳来对DNA片段进行观察,凝胶电泳是根据片段的长度来对片段进行分离。
可以使用连接酶将DNA片段连接在一起。通过将不同来源的DNA片段结合(“连接”)在一起,研究人员可以创造出重组DNA ,这种DNA通常与转基因生物相关。重组DNA技术通常用于质粒:含有一些基因的短环状DNA分子。在分子克隆的过程中,研究人员可以通过将质粒导入细菌中,然后在琼脂平板上扩增DNA片段(分离细菌的克隆——“克隆”也可以指创建克隆(“克隆”)生物体的各种方法)。
DNA也可通过聚合酶链式反应来进行扩增。通过使用特定的DNA短片段,PCR可以分离并指数扩增目标DNA。因为PCR技术可以对含量极少量DNA片段进行扩增,所以PCR也经常用来检测特定DNA序列是否存在。
DNA测序和基因组学
DNA测序是进行遗传学研究的常用方法之一,它可以帮助研究人员测定DNA片段中的核苷酸序列。由弗雷德里克·桑格领导的团队于1977年开发的链终止测序技术,至今仍然常用于对DNA片段进行测序。凭借这项技术,研究人员能够对许多与人类疾病相关的分子序列进行研究。
随着测序成本越来越低,研究人员使用一种叫做基因组组装的方法对多种生物体进行基因组测序,这种方法利用计算工具将许多不同片段的序列拼接在一起。这些技术在2003年完成的人类基因组计划中用于对人类基因组进行测序。新的高通量测序技术大大降低了DNA测序的成本,许多研究人员希望能将人类基因组测序的成本降低到1000美元以下 。
正是对低成本测序的需求不断增加,促进了第二代DNA测序技术(或高通量测序)的出现。这些测序技术允许同时产生数百万个潜在序列。大量可用的序列数据导致基因组学的产生,开创了利用计算工具来搜索和分析生物全基因组的模式。基因组学也可以被认为是生物信息学的一个分支,它使用计算方法对大量的生物数据进行分析。这些研究领域的一个共同点是如何管理和共享涉及人类主题和个人身份信息的数据。另请参见基因组数据共享。
社会和文化
2015年3月19日,一群顶尖生物学家敦促在全球范围内禁止在临床上以可遗传的方式对人类的基因组进行编辑,特别是使用CRISPR和锌指这些方式。2015年4月,中国研究人员展示了利用CRISPR技术对DNA进行编辑的人类胚胎不能存活的研究。
本文转载于:搜狗科学,baike.sogou.com/kexue/d11183.htm,本内容使用CC-BY-SA 3.0授权,用户转载请注明出处