感光细胞是什么?关于感光细胞的科普介绍

感光细胞是在视网膜中发现的一种特殊的神经上皮细胞,能够进行视觉光传导。光感受器最重要的生物学意义在于,它们将光(可见电磁辐射)转化为能够激发生物过程的信号。这个过程更具体是指,细胞中的感光蛋白吸收光子,触发细胞膜电位的变化。
目前已发现哺乳动物眼睛中存在三种不同类型的感光细胞:杆状、圆锥状和固有感光视网膜神经节细胞。最经典的两种感光细胞是杆状细胞和圆锥状细胞,每种都提供信息给视觉系统用来形成视觉世界的展示景象。杆状细胞比视锥细胞窄,虽然两种细胞在视网膜上分布不同,但是用来进行光传导的化学过程是相似的。 在20世纪90年代发现了第三类哺乳动物感光细胞: 固有感光视网膜神经节细胞。这些细胞并不直接提供视觉信息,但被认为在昼夜节律和瞳孔反射中起作用。
杆状细胞和圆锥状细胞的功能存在很大的差异。杆状细胞非常敏感,可以由一个光子触发。 在非常低的光照水平下,视觉信息完全由杆状细胞提供。
圆锥状细胞需要明显更亮的光(即更多的光子)来产生信号。人类有三种不同类型的锥形细胞,它们对不同波长光的反应模式不同。色彩体验主要产生于这三种不同的信号,或许是通过对立过程实现。 这解释了为什么在弱光下当只有杆状感光细胞活跃,而锥形感光细胞处于静置状态时不能分辨出颜色。这三种类型的锥形细胞对短、中和长波长(大致的)的光有反应,因此它们可以分别被称为S-锥体、M-锥体和L-锥体。根据单变量原则,细胞的激活只取决于吸收的光子数量。三种类型的锥形细胞的不同反应可能是由它们各自的感光蛋白吸收不同波长光子所决定。例如,一个L锥形细胞含有一种感光蛋白,这种蛋白更容易吸收长波长的光(即更“红色”)。较短波长的光也能产生相同的响应,但要做到这一点,它必须要亮得多。
人类视网膜包含大约1.2亿个杆状细胞和600万个锥形细胞。根据动物主要是白天活动还是夜间活动的不同,杆状细胞与圆锥细胞的数量和比例也存在差异。某些猫头鹰,如夜间动物灰林鸮, 视网膜中有大量的杆状细胞。在人类视觉系统中,除了感光杆状细胞和圆锥状细胞,还有大约240万到300万个神经节细胞,这些神经节细胞中1%到2%是感光的。神经节细胞的轴突形成两个视神经。
感光细胞通常排列成不规则但近似六边形的网格,称为视网膜镶嵌。
松果体和副松果体在非哺乳动物脊椎动物中是感光的,但在哺乳动物中并不感光。鸟类心室旁器官中有光活性脑脊髓液接触神经元,在对眼睛或神经递质缺乏刺激的情况下对光做出反应。 昆虫和软体动物等无脊椎生物中的光感受器在形态组织和潜在的生化途径上都是不同的。这篇文章只对人类的光感受器进行描述。
组织学
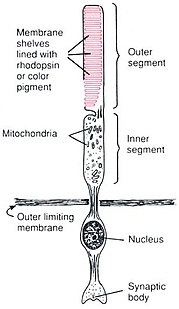
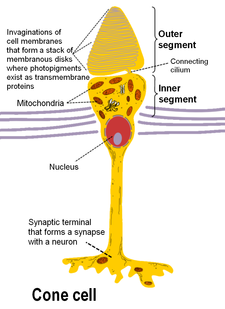
Anatomy of rods and cones varies slightly.
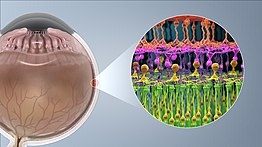
杆状和锥形光感受器位于视网膜的最外层;它们具有相同的基本结构。轴突末端离视野最近(也离大脑最远),可以向双极细胞释放一种叫做谷氨酸盐的神经递质。更靠后的是包含细胞器的细胞体。更靠后的是内部部分,这是细胞中充满线粒体的特殊部分。内部部分的主要功能是为钠钾泵提供三磷酸腺苷(能量)。最后,离大脑最近(也离视野最远)的是外层,即感光体吸收光的部分。外部节段实际上是经修饰的纤毛 包含填充有视蛋白(一种吸收光子的分子,也是电压门控钠通道)的圆盘。
膜感光蛋白opsin含有一种叫做视黄醛的色素分子。在杆状细胞中,这些色素分子统称为视紫红质。在锥形细胞中,有不同类型的视蛋白与视网膜结合形成被称为光蛋白的色素。锥形细胞中三种不同种类的光蛋白分别对不同范围的光频率做出反应,这种差异使得视觉系统能够产生颜色。感光细胞的功能是将光子的光能转换成一种能在神经系统进行传递并能够被生物体所利用的能量形式:这种转换被称为信号转导。
在视网膜固有光敏神经节细胞中发现的视蛋白被称为黑视蛋白。这些细胞参与大脑和身体对(白天)光线的各种反射反应,例如昼夜节律的调节、瞳孔反射和其他对光线的非视觉反应。黑视蛋白的功能类似于无脊椎动物的视蛋白。
当光激活黑视蛋白信号系统时,含有黑视蛋白的神经节细胞产生神经冲动,这些神经冲动通过轴突传导到大脑的特定区域。这些目标包括橄榄前视核(负责控制瞳孔的中心)、LGN,以及通过视黄质下丘脑束的视交叉上核(RHT,昼夜节律的主起搏器)。含黑视蛋白的神经节细胞被认为通过从轴突末端释放神经递质谷氨酸盐和垂体腺苷酸环化酶激活多肽(PACAP)来对这些目标产生影响。
人类

人类视网膜大约有600万个视锥细胞和1.2亿个视杆细胞。来自杆状细胞和圆锥细胞的信号汇聚在神经节细胞和双极细胞上进行预处理,然后发送到外侧膝状体核。视网膜的“中心”(透镜正后方的点)是小凹(或中央凹),它只包含锥形细胞;它是能够产生最高视觉灵敏度或最高分辨率的区域。在视网膜的其余部分,杆状细胞和圆锥状细胞混合在一起交错分布。盲点中没有发现光感受器的存在,在盲点中,神经节细胞纤维组织成视神经并离开眼睛。
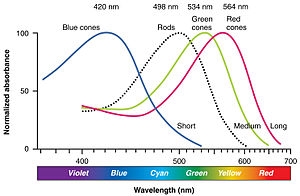
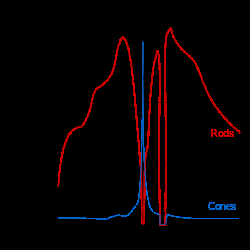
三种类型锥体中的感光蛋白对不同波长光子的敏感度不同(见图)。因为视锥细胞对光的波长和强度都有响应,如果刺激的强度保持不变,而当波长变化时,锥体对波长的灵敏度是根据其相对响应速率来进行评估的。由此反过来,可以推断吸光度。 该图在百分制图表上将吸光度标准化。例如,视锥细胞的相对响应峰值约为420纳米(纳米,波长的度量单位)。这告诉我们,与任何其他波长相比,S视锥细胞更倾向于吸收420纳米的光子。然而,如果是不太敏感的不同波长的光,例如480纳米,适当增加亮度,它将在S视锥细胞中产生完全相同的响应。所以,曲线的颜色是误导性的。视锥细胞本身无法检测颜色;相反,色觉需要对不同圆锥类型的信号进行比较才能产生。
光转换
光传导的过程发生在视网膜上。 视网膜有许多不同细胞类型的层。 最著名的感光细胞(杆状和锥形)形成最外层。他们是负责视觉的光感受器。中间层包含双极细胞,双极细胞从杆状细胞和视锥细胞收集神经信号,然后将它们传输到视网膜最内层, 在那里的神经元被称为视网膜神经节细胞(RGCs)(其中一小部分本身是光敏的),组织信号并将其发送到大脑。
杆状细胞和圆锥细胞的激活实际上是超极化;当它们处于静止状态时,它们去极化并持续释放谷氨酸盐。在黑暗环境中,细胞具有相对高浓度的环鸟苷3’-5’单磷酸(cGMP),它打开离子通道(主要是钠通道,钙也可以通过这些通道进入)。沿着电化学梯度进入细胞的正电荷离子改变了细胞的膜电位,引起去极化,并导致神经递质谷氨酸的释放。谷氨酸能使一些神经元去极化,并使其他神经元超极化。
当光照射感光细胞内的感光色素时,色素的构象会发生改变。这种色素被称为视紫蓝质或视紫红质,由一种叫做视蛋白(位于质膜中)的大蛋白质组成,附着在共价结合的辅基上:一种叫做视黄醛的有机分子(维生素A的衍生物)。视黄醛在黑暗中以11-顺式视黄醛的形式存在,光的刺激使它的结构发生改变成为反式视黄醛。这种结构变化导致它激活一种被称为转导素的调节蛋白,从而导致cGMP磷酸二酯酶的激活,该酶将cGMP分解为5'-GMP。cGMP的减少使离子通道关闭,阻止正离子的流入,使细胞超极化,并阻止神经递质的释放。光引发感官反应的整个过程称为视觉光传导。
暗电流
未被刺激的环境中(在黑暗中),因为环GMP与外部片段的环核苷酸门控通道结合,所以门控通道是开放的。因此,带正电荷的离子(即钠离子)进入感光体,使其去极化至约40毫伏(其他神经细胞的静息电位通常为65毫伏)。这种去极化电流通常被称为暗电流。
信号转导途径
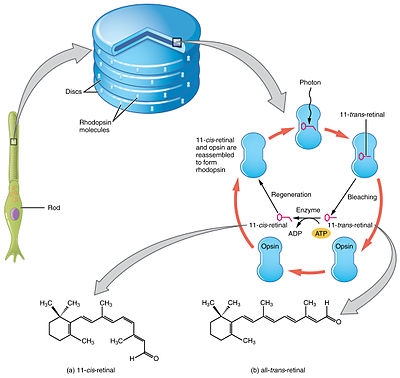
信号转导途径是光子能量向细胞中传导从而导致其极化进而产生信号的机制。这种极化最终导致那些通过视神经输入大脑神经信号被传输或被抑制。信号在脊椎动物眼杆和视锥光感受器中转导步骤或转导途径是:
1、外节盘膜中的视紫红质或视紫蓝质吸收光子,将蛋白质内视网膜希夫碱辅因子的构型从顺式改变为反式,从而导致视网膜的形状发生改变。
2、这些变化会导致一系列不稳定中间体的产生,最后一个中间体与膜上的一种被称为转导素的G蛋白以更强的化学键相结合并将其激活。这是扩增反应的第一步——每个光激活的视紫红质触发大约100个转导素的激活。
3、然后每个转导素激活cGMP特异性磷酸二酯酶(PDE)。
4. 然后PDE催化cGMP水解成5'-GMP。这是扩增的第二步,其中每个PDE水解约1000个cGMP分子。
5. 细胞内cGMP的净浓度降低(由于它被PDE转化为5'-GMP),导致位于感光细胞外节膜的环核苷酸门控钠离子通道关闭。
6. 最终导致钠离子不再能进入细胞,从而使膜内的电荷变得更负,导致感光体外膜变得超极化。
7. 细胞膜电位的这种变化导致电压门控钙通道关闭。这导致流入细胞的钙离子减少,因此细胞内钙离子浓度下降。
8. 细胞内钙浓度的降低意味着通过钙诱导的胞吐作用向双极细胞释放的谷氨酸盐减少(见下文)。(细胞内钙离子浓度的降低,从而减缓了用以刺激位于突触后的双极细胞和水平细胞的神经递质谷氨酸的释放。)
9. 谷氨酸释放的减少意味着一群双极细胞将去极化,另一群双极细胞将超极化,这取决于突触后最终受体(离子型或代谢型)的性质(见感受场)。
因此,当受光刺激时,杆状或圆锥状感光体实际上释放较少的神经递质。感光细胞和双极细胞之间突触间隙中较少的神经递质将会激活双极细胞(去极化)或抑制双极细胞(超极化)。因此,在光感受器-双极细胞突触处,视觉信号被分成开和关通路。
内部提供的三磷酸腺苷为钠钾泵提供动力。这种泵对于来重置外层的初始状态很有必要,作用原理是通过吸收进入细胞的钠离子并将其泵出。
尽管光感受器是神经元,但除了光敏神经节细胞外,它们不传导动作电位——光敏神经节细胞主要参与昼夜节律、褪黑激素和瞳孔扩张的调节。
优势
杆状细胞和锥状细胞的光传导有些不寻常,因为刺激(在这种情况下是光)会降低细胞的反应或激发率,不同于大多数其他感觉系统中刺激会增加细胞的反应或激发率。这种差异具有重要的功能性影响:
首先,经典(棒状或锥形)感光体在黑暗中去极化,这意味着有更多的钠离子可以流入细胞。钠通道的随机打开或关闭并不会对细胞的膜电位产生影响;只有通过吸收光子,从而导致大量通道关闭,才会对膜电位产生影响产生光在视野中的信号。相对于感觉传导模式,该系统的噪声更小,感觉传导模式相较于神经放电模式来说,对刺激(如触摸和嗅觉)的反应速度更快。
第二,在经典光转化的两个阶段中,每一步对所产生的结果均进行放大:一种色素将激活许多转导素分子,而一种PDE将会催化很多cGMPs发生裂解。这种放大意味着即使只吸收到一个光子也会对膜电位产生影响,并向大脑发出光在视野中的信号。这是棒形感光体和锥形感光体的主要区别。与锥形感光体不同,棒形感光体非常敏感,有记录单个光子的能力。另一方面,视锥细胞与杆细胞不同,视锥细胞可以很快的对光传导信号进行放大。
视锥细胞与视杆细胞的区别
艾瑞克·坎德尔等人在《神经科学原理》中对人类杆细胞和圆锥细胞的比较。
| Rods | Cones |
|---|---|
| Used for scotopic vision (vision under low light conditions) | Used for photopic vision (vision under high light conditions) |
| Very light sensitive; sensitive to scattered light | Not very light sensitive; sensitive to only direct light |
| Loss causes night blindness | Loss causes legal blindness |
| Low visual acuity | High visual acuity; better spatial resolution |
| Not present in fovea | Concentrated in fovea |
| Slow response to light, stimuli added over time | Fast response to light, can perceive more rapid changes in stimuli |
| Have more pigment than cones, so can detect lower light levels | Have less pigment than rods, require more light to detect images |
| Stacks of membrane-enclosed disks are unattached to cell membrane directly | Disks are attached to outer membrane |
| About 120 million rods distributed around the retina | About 6 million cones distributed in each retina |
| One type of photosensitive pigment | Three types of photosensitive pigment in humans |
| Confer achromatic vision | Confer color vision |
功能
光感受器不发出色彩信号;它们只表示视野中有光。
感光体对光源的波长和强度都可以产生响应。例如,特定强度的红光可以在感光体中产生与某个强度的绿光产生相同的反应。因此,单个感光器对颜色进行分辨所产生的结果是不准确的。
发展
诱导杆状细胞、S型锥状细胞、M型锥状细胞分化的关键是由几种转录因子所介导的,包括RORbeta、OTX2、NRL、CRX、NR2E3和TRbeta2。S型视锥发育代表默认的感光细胞程序,然而不同的转录活性可以导致杆状或M型视锥的产生。L锥体存在于灵长类动物中,但是由于在研究中使用啮齿动物,目前并不清楚L型椎体的发育过程。光感受器的产生分为五个步骤:多能视网膜祖细胞的增殖;抑制视网膜祖细胞;细胞分化;感光细胞基因表达;最后是轴突生长、突触形成和外节生长。
早期Notch信号维持祖细胞循环。光感受器前体是通过抑制 Notch信号传导和增加各种因子的活性而产生的,包括盾片基因同源蛋白 1。OTX2分子活动刺激细胞从而决定光感受器的命运。CRX进一步限定了感光细胞特异性基因组的表达。NRL的表达决定了杆状视觉细胞的发育。NR2E3通过抑制锥体视觉基因的表达进一步促进细胞向杆状视觉细胞分化。杆状细胞和锥状细胞的发育都需要RORbeta。trβ2介导了M锥的命运。如果前面提到的任何因素的功能缺失,默认发育成的感光器是一个S型视锥细胞。这些事件发生在不同物种的不同时期,是一个复杂的模型,最终导致产生一系列不同的表型。如果这些调节网络被破坏,可能导致色素性视网膜炎、黄斑变性或其他视觉缺陷。
信号传递
杆状和锥形光感受器通过减少轴突末端分泌神经递质谷氨酸到的双极细胞来发出吸收光子的信号。由于感光细胞在黑暗中去极化,大量谷氨酸盐在黑暗中被释放到双极细胞中。吸收的光子会使光感受器超极化,因此导致突触前末端向双极细胞释放较少的谷氨酸盐。
每个杆状或锥形感光细胞释放相同的神经递质谷氨酸盐。然而,由于位于细胞膜的受体类型不同,谷氨酸盐在双极细胞中的作用也是不同的。当谷氨酸盐与离子型受体结合时,双极细胞将去极化(因此,随着谷氨酸盐释放的减少,双极细胞将被光超极化)。另一方面,谷氨酸与代谢型受体的结合导致受体超极化,因此当谷氨酸释放减少时,双极细胞将因为光去极化。
本质上,即使所有的光感受器对光都可以产生相同的反应,但上文提到的这种特性可以允许一个双极细胞群体因受到光影响而被激发,而另一个群体因为光的刺激而被抑制。在检测颜色、对比度、边缘等特征时这种复杂的反应是十分重要且不可或缺的。
进一步的复杂性来自视网膜中双极细胞、水平细胞和无轴突细胞之间的各种相互联系。最终结果是视网膜中神经节细胞分化为不同群体,其中存在某些含有感光色素黑色素具有感光性的亚群。
神经节细胞(非杆状或锥形感光细胞)光感受器
福斯特(Foster)等人 于1991年发现了小鼠眼中的一种非杆状非锥体感光细胞,它被证明能调节昼夜节律,这些神经元细胞被称为固有光敏视网膜神经节细胞(ipRGC),是一小部分(约1-3%)位于内视网膜的视网膜神经节细胞, 即位于外视网膜杆状细胞和锥形细胞的前面。这些光敏神经元含有一种光色素——黑视蛋白,它的光吸收峰与杆状细胞和圆锥细胞的波长不同(约480纳米)。除了昼夜节律/行为功能外,ipRGCs在启动瞳孔光反应方面也有作用。
丹尼斯·戴西和他的同事在一种旧大陆猴身上发现了一种可以产生投射到外侧膝状体核(LGN)的视黑素的巨大神经节细胞。 以前只发现了中脑(顶盖前核)和下丘脑(视交叉上核)的投射。然而,受体的视觉作用仍未有确切结论。
2007年,法尔汉·扎伊迪和他的同事首次发现了非杆状或锥形的人类感光细胞。随后在2008年《当代生物学》给科学家和眼科医生的社论宣布,他们发现人类非杆状或锥形感光细胞所使用的实验方法,在应用于在其他哺乳动物时,同样可以发现非杆状或锥形感光细胞的存在, 人类的无杆锥状感光细胞的本质是内部视网膜中的神经节细胞。工作人员已经发现存在缺失传统的杆状和圆锥状感光细胞功能,但保留了神经节细胞功能的罕见疾病的患者。 尽管没有杆状细胞或圆锥状细胞,但患者仍然存在昼夜光调节、昼夜行为模式、黑视蛋白抑制和瞳孔反应,这些表现与环境和实验光的峰值光谱灵敏度与黑视蛋白光调节相匹配。他们的大脑也能把视觉和这种频率的光联系起来。
在人类中,视网膜神经节细胞感光细胞对有意识的视觉以及非成像功能,如昼夜节律、行为和瞳孔反应都有一定作用。 因为这些细胞主要对蓝光有反应,所以有人认为它们在中间视觉中有作用。Zaidi及其同事对无杆锥状人类受试者的研究,因此也为神经节细胞光感受器的成像(视觉)所起的作用打开了大门。人们发现有平行的视觉通路——一个来自外层视网膜的典型的杆状和圆锥状视觉通路,另一个基本的视觉亮度检测器通路来自内层视网膜,它似乎在另一个之前被光激活。 经典的感光体也为新型的光感系统提供了新的见解,福斯特发现色觉恒定性可能在新的感光体系统有重要作用。这种受体有助于理解许多疾病,包括全球性的失明的主要原因,如青光眼,这是一种影响神经节细胞的疾病,对这种受体的研究为寻找治疗失明的新方法提供了可能。尽管昼夜节律紊乱的影响是与临床医学相关的另一个领域,正是这些对人类新感光体的发现和受体在视觉中的作用进行研究,而不是研究它的非成像功能,对受体进行研究可能对整个领域产生巨大的影响。
大多数工作表明,受体的峰值光谱灵敏度在460和482纳米之间。Steven Lockley等人在2003年指出,460纳米波长的光抑制褪黑激素的能力是555纳米波长光的两倍。然而,在Farhan Zaidi等人最近的工作中,发现人类非杆状或锥形感光细胞在481纳米的刺激下对光的感觉能力是最强的;这意味着,视觉受体的最大限度是在蓝光条件下提供一些基本的视觉。
本文转载于:搜狗科学,baike.sogou.com/kexue/d10036.htm,本内容使用CC-BY-SA 3.0授权,用户转载请注明出处