原核生物的转录后加工:
mrna转录后加工:少数多顺反子mRNA需要由核酸内切酶切成较小的单位,再进行翻译,主要是有利于分别进行调控少数噬菌体(如T4)中发现由内含子,需要拼接。
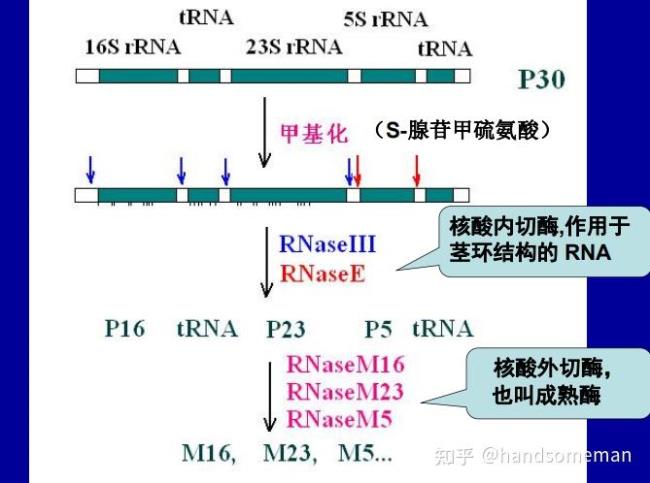
rRNA的转录后加工:E.coli.rRNA gene 与某些tRNA gene组成共转录单位(rrn operons),一共有7个这样的转录单位共转录物经剪切、修剪和修饰后得到成熟的rRNA和tRNA。
tRNA的转录后加工:E.Coli中多数tRNA基因分散存在于基因组DNA中,少数于rRNA基因共转录初级转录物两侧都有多余序列,需要经历转录后加工加工方式包括剪切、修建和核苷酸修饰。
过程:核酸内切酶切去前体两端序列:RNase P产生成熟5'端RNaseF切去3'端部分附加序列核酸外切酶(RNaseD)从3'端逐个切去其余附加序列,产生成熟的3'端核苷的修饰(碱基)。
RNase P,RNaseF,RNaseIII,RNaseE都是核酸内切酶,但是与DNA限制性内切酶不同,不是识别特定的核苷酸序列,而是识别特定的空间结构。
RNaseP是个特殊的酶:RNase和蛋白酶处理都可以使RNaseP失活特定条件下,M1RNA可以单独切断tRNA的5'末端。
二、真核生物的转录后加工
1、真核生物转录后加工的特征
mRNA:转录和翻译再时间和空间上都被分开,所以转录加工十分重要
tRNA,rRNA:需要转录后加工
2、rRNA的转录后加工
rRNA基因是串联排列的中度重复序列除5SrRNA外的其他rRNA作为一个转录单位转录核仁是rRNA转录、加工并装配成核糖体的场所主要过程与原核生物rRNA前体加工相似依赖于RNaseIII和其它核酸内切酶甲基化程度更高需要snoRNA的帮助有些生物的rRNA前体含有内含子,需要拼接
哺乳动物的18S,28S,5.8SrRNA gene组成一个转录单位,有RNApolI转录产生45S的前体分子5SrRNAgene与不同转录区域组成转录单位,由RNA pol III转录
small nucleolar RNA(snoRNA):有几百种snoRNA,但只有少数参与rRNA前体加工snoRNA与特定蛋白质组装成snoRNP后才能起作用snoRNP中的RNA可以rRNA前提中的特定序列互补配对,以确定修饰位点
3、tRNA的转录后加工:主要过程与原核生物tRNA前体加工相似,tRNAgene成簇排列,并被间隔区分开,由RNA pol III转录需要多种核酸内切酶和外切酶核内RNase P的RNA部分没有活性细胞器中无RNase P前体的3'端没有CCA序列有些有内含子。因此加工方式包括:剪切、修剪、碱基修斯、添加CC、拼接添加CCA和拼接是真核生物特有的两种后加工方式。
4、mRNA的转录后加工
真核生物中编码蛋白质的基因转录产物为单顺反子mRNAmRNA的原初转录物是分子量极大的前体,在核内加工过程中形成分子大小不等的中间物,被称为核内不均一RNA(heterogeneous nuclear RNA,hnRNA)
加工过程(processing):5'端加帽(capping)3'端加尾(tailing)内部甲基化(internal methylation)拼接(splicing)编辑(editing)
4、1 5'端加帽:
绝大多数真核生物的细胞核mRNA和某些snRNA的5'端含有帽子结构帽子有0型、1型、2型三种酵母mRNA的帽子为0型脊椎动物mRNA的帽子为2型snRNA的帽子结构比较特殊
加帽反应为共转录反应
一些snRNA带有其他类型的帽子结构:2,2,7-trimethylated guanosine
帽子结构的功能:保护mRNA,防止其被讲解增强mRNA的可翻译性促进成熟mRNA从细胞核转运到细胞质中促进mRNA的拼接(有助于去除第一个内含子)。
mRNA加帽原因:只有RNA聚合酶II合成的转录产物(mRNA、部分snRNA)才有帽子结构因为加帽酶只能与RNA聚合酶II的CTD结构域结合而CTD是RNA聚合酶II特有的加帽酶和CTD的磷酸化形式(延申型)结合转录产物一旦从RNA聚合酶II中显露出来,就可以与加帽酶接触。
4、2 3'端加尾
真核生物的大多数mRNA及其前体在3'端有约250nt的连续AMPpoly(A)由poly(A) polymerase(PAP)添加mRNA进入细胞质后,其poly(A)可以被更新不断得被RNase降解,再由细胞质中的PAP重新合成。真核生物mRNA的3'端都有polyA,但组蛋白mRNA没有polyA加尾过程在核内已经完成,核内hnRNA也有polyA。
polyadenylation signal(加尾信号)是一种顺式作用元件:真核生物(酵母除外)和病毒mRNA的3'端附近有一段保守序列AAUAAA(最常见,有效的加尾信号,AUUAAA大约有80%的有效性,其他变体既不常见,也不有效),称为加尾信号,删除后不能加尾加尾信号在下游20nt左右进行多聚腺苷酸化。
其他顺式元件:动物细胞还包括:GU-rich motif和U-rich motif。植物细胞还需要U-rich motif酵母细胞的加尾元件差别较大。
加尾所需的蛋白质因子(反式因子):剪切与多聚腺苷酸化特异性因子(CPSF)特异性结合AAUAAAsignal剪切刺激因子(CstF):结合GUregion、与CPSF结合剪切因子I和II(CFI、CFII):核酸内切酶poly(A)聚合酶(PAP)PNA pol IIpoly(A)结合蛋白(PABP)
加尾过程:第一阶段:CPSF识别 AAUAAA信号,招募 CF I、CF II、CstF、PAP等。CF I 和 II在下游 切断RNA,PAP添加约 10个AMP(慢反应)第二阶段:PABP与已形成的polyA结合, PABP、CstF使反应加速, PAP继续添加200个 以上的 AMP(快反应)。
尾巴的功能:保护mRNA防止降解促进mRNA翻译促进mRNA从细胞核转运到细胞质提高mRNA的拼接效率(最后一个内含子的去除)产生终止密码子通过选择性加尾调节基因表达。
mRNA加尾巴原因:跟加帽类似
4、3 内部甲基化
主要是m6A在hnRNA已经存在功能不清楚
4、4 拼接(splicing)
大多数真核生物基因是断裂基因
其中编码序列称为外显子(exon),外显子之间的介入序列称为内含子(intron)少数真核生物基因(如组蛋白、干扰素)是连续的高等真核生物的基因中多数内含子比外显子长得多,而低等真核生物(如酵母)的基因中内含子比较短而且少见含内含子最多基因:人巨肌蛋白(titin)基因,362个内含子。一些tRNA基因、rRNA基因也有内含子tRNA基因的内含子较小(4-50bp)线粒体、叶绿体基因也有内含子。
RNA拼接定义:去除内含子,连接外显子称为成熟mRNA的过程称为RNA拼接。
I类内含子拼接:多数真核生物rRNA基因不含内含子少数含内含子但不转录(如果蝇1/3rRNA基因含内含子但不转录)四膜虫(tetrahymena)的核rRNA基因、酵母线粒体rRNA基因含内含子,并转录
四膜虫rRNA基因的拼接:需要G(GTP,GDP,GMP,G)不需要任何蛋白质的参与称为自我拼接self-splicing或者自身催化auto-catalysis分步很广,存在于低等真核生物rRNA基因、真核生物细胞器基因、细菌和噬菌体个别基因I类内含子内含子部分(L19 IVS)与传统酶有许多共性:提高反应速度、专一性等但是反应前后自身发生变化,所以不是严格意义上的酶。但经实验证明他是酶。
核酶(ribozymes):具有催化活性的RNARNase P中的M1RNAL19 IVS.
II 类内含子的拼接:酵母Cyt C氧化酶亚基a基因的拼接:内含子3'拼接点上有一个百分之百保守的A也属于自我拼接不需要鸟苷或鸟苷酸存在于某些真菌线粒体和植物叶绿体基因。
I、II型内含子都能形成特征性的二级结构都可以自我拼接都可以作为转座子:I型内含子通过DNA介导的机制而移动,II型内含子通过RNA介导的机制而移动。I型编码核酸内切酶,II型编码逆转录酶。
真核生物核内pre-mRNA内含子的拼接:高度准确:依赖于多种顺式作用元件和反式作用因子共转录事件(转录还没有结束时候就开始了)
顺式元件1:内含子具有一致的保守序列,即5'拼接点为GU,3'拼接点为AG,称为GU/GT-AG规则。
顺式元件2:3'拼接点上游的一段富含嘧啶的序列约11个碱基组成靠近3'拼接点。
顺式元件3:内含子序列内部有一个保守序列:分支点(branch point)。分支点保守序列决定下游的AG为3'拼接点。
反式因子:拼接反应在拼接体spliceosome中进行,酵母的拼接体约40s,哺乳动物的拼接体约60s多种反式因子参与的复杂过程。
snRNP:small nuclear ribonucleoprotein particlesrecognize the splicing signal